Los reinos de la vida (página 5)
Euglena sea común protists, de la clase Euglenoidea de phylum Euglenophyta. Actualmente, sobre 1000 especies de Euglena se han descrito. Marin y otros. (cloroplastos, clasificado antes como Astasia y Khawkinea. El Euglena puede ser considerado a veces para tener características de la planta y del animal.
Un euglena es un protist que puede comer el alimento como lo hacen los animales (en parte heterotrophic) y puede hacer el alimento como lo hacen las plantas (en parte autotrophic). El Euglena puede rodear una partícula del alimento y comerla (fagocitosis). Mucho el Euglena contiene los cloroplastos y la clorofila a y B. Euglena vivo en el agua dulce, agua salada y en el suelo. Mucho el Euglena puede moverse usando un flagelo, un largo azotar-como la estructura. Cuando el agua se seca para arriba, las formas de un euglena una pared protectora gruesa alrededor de sí mismo y las mentiras inactivas bajo la forma de espora hasta que el ambiente mejora. Se reproduce por la fisión como ameba.
Alimentación: La obtienen principalmente por fotosíntesis en proceso interno (nutrición holofítica). Sin embargo, necesitan materiales como la vitamina B12, etc., que no pueden producir pero que, al igual que varios minerales, pueden absorber del medio circundante. En la oscuridad, pueden vivir a expensas de compuestosorgánicos disueltos en el agua (nutrición saprófita). Bajo estas condiciones, los cloroplastos y los pirenoides desaparecen.

Estructura de una euglena:
protozoo flagelado que vive en las aguas dulces. Es compuesto de clorofilo y de un ojo rudimentario.
Reservorio: parte de una euglena que sirve a acumular.
Nucleo: orgánulo central de una euglena.
Vacuola contráctil: cavidad de una euglena que tiene la capacidad de contraerse.
Membrana celular: membrana que envuelve euglena.
Cloroplasta: orgánulo de una euglena donde es la fotosintésis.
Nucléolo: cuerpo con forma de una esfera en el nucleo de una euglena.
Estigma: parte de una euglena sensible a la luz.
Flagelo: filamento móvil y largo que sirve a la locomoción de una euglena.
CAPITULO III:
Las traqueofitas
(Phylum: Tracheophyta)
Tracheobionta es el nombre del taxón de plantas que abarca a las traqueófitas, también llamadas plantas vasculares. Son organismos formados por células vegetales, que poseen un ciclo de vida en el que se alternan las generaciones gametofítica y esporofítica, siendo esta última la fase dominante (sobre quien actúa más presión de selección natural); cuya fase esporofítica es fotosintética e independiente, y tiene tejidos y sistemas de órganos; está organizada en un "cormo" (sistema que posee vástago aéreo, raíz subterránea y un sistema de conducción vascular que los vincula) que es a lo que comúnmente se refiere la gente cuando dice "planta"; cuya fase gametofítica es reducida y puede ser desde un "talo" (cuerpo no organizado en tejidos ni órganos) en helechos y afines, hasta unas pocas células protegidas y nutridas por el esporófito, en gimnospermas y angiospermas. La selección natural dirigió fuertemente la evolución de las traqueófitas hacia una menor dependencia de las condiciones ambientales sobre la tierra para la reproducción y la dispersión, característica que entra en evidencia al comparar las traqueofitas más antiguas (Lycophyta) con las más modernas (plantas con flores).
A lo largo de la historia se ha designado a este grupo con distintos nombres, como cormófitas o cormofitas (nombre científico Cormophyta o Cormobionta), embriófitas sifonógamas (nombre científico Embryophyta sifonogama), etc. Su nombre más común, Tracheophyta, proviene de tracheo (en referencia a las traqueidas, especializadas para el transporte de líquidos dentro de la planta) y phyta, raíz de origen griego que significa "planta". Se lo castellaniza como traqueofitas o traqueófitas.
La evidencia de los análisis moleculares de ADN hoy en día demostró que las traqueófitas son un grupo monofilético (que comprende a todos los descendientes de un ancestro común) dentro de las embriófitas, mientras que las briofitas son parafiléticas con respecto a las traqueofitas. Esto quiere decir que probablemente las traqueófitas sean descendientes de plantas muy parecidas a los briófitos, con el gametofito siendo la fase dominante, y el esporofito sin ramificar y dependiente nutricionalmente del gametofito.
Dentro de las traqueófitas hay dos linajes principales, Lycophyta y Euphyllophyta, diferenciadas principalmente por la forma de construcción de sus hojas (en las licofitas son lycofilos y en las eufilofitas son eufilos, los eufilos se corresponden aproximadamente con los megafilos, aunque en algunos grupos pueden estar reducidos en forma secundaria a su adquisición). Las eufilofitas a su vez comprenden dos grandes linajes, Monilophyta (helechos, equisetáceas y psilotáceas) y Spermatophyta, que se diferencian entre sí porque el primero tiene gametofitos de vida libre y el segundo los tiene encerrados en la semilla y el grano de polen. A su vez las espermatofitas están formadas por dos grandes grupos monofiléticos vivientes, Gymnospermae y Angiospermae o Magnoliophyta, que se diferencian entre sí porque el primer linaje tiene las semillas a la vista sobre la hoja fértil, mientras que el segundo tiene las semillas encerradas dentro de las paredes de la hoja fértil o carpelo.
Aún se siguen estudiando a las Lycophyta y Monilophyta como agrupadas en el grupo parafilético de los "helechos y afines" o Pteridophyta. Las traqueofitas son un grupo importante tanto por dominar la mayor parte de los ecosistemas terrestres como por ser muy utilizadas por el hombre.
El esporofito de las plantas vasculares:
El cuerpo vegetativo del esporófito es un "cormo" (vástago + raíz + sistema de conducción vascular, hecho gracias al engrosamiento de las paredes celulares de células alargadas), que además de los tejidos vasculares posee tejidos de protección y tejidos de sostén, y que crece gracias a la acción de sus meristemas anatomy/11.html anatomy/14.html anatomy/13.html.
El "cormo" está especializado para la vida terrestre. Consta de vástago, raíz y un sistema de conducción vascular que los vincula. El vástago es la región del cormo que se especializa en la fotosíntesis, y que utiliza para ello el agua y las sales minerales que le llegan de la raíz. La raíz es la región del cormo que se especializa en la absorción de agua y sales, y que utiliza para ello los azúcares proporcionados por el vástago. Esta especialización es posible gracias a que el cormo posee un sistema de conducción vascular formado gracias a que las paredes de la célula vegetal pueden volverse ricas en lignina, compuesto que les otorga dureza.
Meristemas:
El cormo crece gracias a la actividad de sus meristemas (del griego meros: dividir) que son un grupo de células en estado embrionario permanente, capaces de dividirse indefinidamente, formando tejidos que en su juventud son indiferenciados anatomy/11.html anatomy/14.html anatomy/13.html .
Los principales tipos de meristemas son dos:
en todos los cormos encontraremos en los ápices de los tallos y las raíces, meristemas primarios responsables del crecimiento primario del esporofito.
en los cormos con crecimiento secundario del tallo, encontraremos meristemas secundarios a lo largo del tallo, encargados de su crecimiento en grosor.
Sistemas de tejidos:
El botánico J. Sachs en el siglo XIX, distinguió tres sistemas principales de tejidos en el cormo del esporofito de las cormófitas, clasificados según su función en la planta: tejidos de protección, tejidos fundamentales, y tejidos vasculares.
Tejidos de protección:
Los tejidos que cumplen la función de protección forman la capa más externa del cormo. Si el cormo sólo posee crecimiento primario, encontraremos una epidermis, cubierta por una capa de cutina (lípido complejo que evita la pérdida de agua en la vida terrestre pero también evita el intercambio gaseoso con el medio ambiente), y con estomas Allies/Psilophyta/Psilotum nudum/Stem xs/Epidermis stomata.html y lenticelas (ambos aseguran el intercambio gaseoso con el ambiente). En los taxones con crecimiento secundario del tallo, la epidermis con cutina es reemplazada durante el crecimiento secundario, por una peridermis parcialmente impermeabilizada con suberina (lípido muy parecido a la cutina, responsable de la formación del súber o corcho).
Tejidos fundamentales:
Los tejidos fundamentales forman un sistema continuo y están conformados principalmente por los diversos tipos de parénquimas (del griego: "carne de las vísceras"), a los que se asocian los tejidos de sostén (como colénquima y esclerénquima).
Los tejidos de sostén, como su nombre lo indica, cumplen la función de mantener la estructura de la planta, función que cumplen gracias a la lignina presente en la pared celular. Los tejidos de sostén más comunes son:
Colénquima (del griego: goma, cola, nombre dado por la facilidad con que las paredes celulares se hinchan al hidratarse) formado por células vivas.
Esclerénquima (del griego escleros: duro, nombre dado por sus gruesas paredes muy duras y resistentes) formado por células casi siempre muertas a su madurez.
Tejidos vasculares:
Son los responsables del transporte de líquidos y sustancias por todo el cuerpo del vegetal, en el que se distinguen el floema (del griego floeos: "yema de la corteza, propio de la corteza", especializado en transporte de azúcares) y el xilema (del griego xylos: "lignificarse", formado por células muertas tubulares, abiertas o cerradas en sus extremos, de paredes muy lignificadas, que forman un haz vascular especializado en transporte de agua y sales) [1] [2] [3] [4] [5]. Estos tejidos son complejos, y a menudo, están asociados a otros (parenquimáticos y de sostén). Los tejidos vasculares se ubican dentro de los tejidos fundamentales de manera diversa según los distintos órganos de la planta (raíz, tallo, etcétera), que en un corte transversal forman patrones (llamados estela), que tienen importancia sistemática. Así el tallo de la mayor parte de las monocotiledóneas presenta un patrón o estela llamado atactostela, el tallo (primario) de las dicotiledóneas y de las coníferas en cambio, presenta una eustela, y el de los helechos y afines presenta una gran diversidad que permite diferenciar a las familias entre sí.
Evolución de traqueófitas:
Mucho de lo que sabemos acerca de las primeras plantas vasculares y su transición a la vida terrestre se encuentra en el registro fósil, ya que esos valiosos linajes están todos extintos.
Las traqueófitas ancestrales derivaron de las primeras embriófitas, las primeras plantas que colonizaron la superficie terrestre. Todo indica que las primeras embriófitas eran pequeñas y muy simples en su estructura. En el caso del linaje que derivó en las plantas vasculares, el esporofito era básicamente un tallo de ramificación dicotómica, al principio de la altura de un fósforo, con el esporangio (donde ocurre la meiosis que da esporas haploides) producido en las puntas de las ramas. Estas plantas no tenían ni hojas ni raíces, en algunos casos (como Rhynia del "Rhynia Chert" en Escocia) la preservación de estas plantas es espectacular, y es posible discernir muchos detalles anatómicos, incluyendo estomas, esporas, y el tejido vascular dentro del tallo.
En los análisis basados en dichos fósiles, fue recientemente descubierto que los primeros polysporangiofitos (plantas con esporofitos ramificados) en realidad no producían verdaderas células conductoras de agua (traqueidas) en el xilema, y por lo tanto deben haber dependido por completo de la presión de turgencia para mantenerse erguidos. Las células verdaderamente conductoras de agua evolucionaron más tarde y caracterizan al clado de las traqueófitas o "verdaderas plantas vasculares" (Kenrick y Crane 1997a,1 1997b2 ).
Las traqueidas son células elongadas con paredes engrosadas, muertas a la madurez. Donde una traqueida se conecta con la siguiente se observan unas aberturas características en la pared secundaria ("pits" en inglés) pero la pared celular primaria de las células que formaron las traqueidas se mantiene intacta, dando la "pit membrane". El agua por lo tanto debe atravesar las paredes primarias de las células para poder avanzar. En las primeras traqueófitas (representadas por Rhynia)las traqueidas era de un tipo distintivo, con cierta "resistencia al desmoronamiento" ("decay resistance"), conferida por la lignificación de las fibras de celulosa, que estaba presente sólo como una capa muy delgada. Las paredes celulares más resistentes son las que caracterizan al clado de las eutracheofitas, que include a todas las plantas vasculares vivientes (Kenrick y Crane 1997a1 ). En estas especies, las traqueidas fuertemente lignificadas permiten una conducción de agua más eficiente, y proveen resistencia interna, permitiendo a las plantas crecer mucho más altas.
Para leer una discusión acerca de la evolución de la predominancia del esporofito en detrimento del gametofito, ver Evolución de embriófitas.
Las relaciones filogenéticas entre las líneas de plantas vasculares vivientes, mostradas en la figura de la sección sistemática, muestran una división basal, que ocurrió en el Devónico temprano a medio (hace unos 400 millones de años), que separa al linaje de las lycophytas actuales del linaje de las eufilofitas actuales. Esta división está marcada por una considerable variedad de características morfológicas. Una bastante notoria es la presencia de esperma multiflagelado en las eufilofitas en oposición al esperma biflagelado de las lycophytas y de los linajes desprendidos de grupos más antiguos (los briófitos sensu lato). Las dos únicas excepciones son Isoetes y Phylloglossum, donde el esperma multiflagelado se originó independientemente.
Las primeras plantas con semilla aparecieron en el Devónico tardío, y eso llevó a la típica flora del Mesozoico dominada por gimnospermas. Los fósiles más antiguos de angiospermas encontrados hasta la fecha datan aproximadamente de hace 140 millones de años (principios del Cretácico), y el número de especies encontrado aumenta de forma llamativa con el tiempo geológico, en comparación a otras plantas no angiospermas (el llamado "abominable misterio" de Darwin). Para una discusión sobre la radiación de las angiospermas ver Evolución de angiospermas.

CLASIFICACION DE LAS TRAQUEOFITAS:
Las Traqueofitas o Plantas Vasculares se Clasifican en:
– Pteridófitas, son Traqueofitas pero Criptógamas ya que sus sexos están ocultos, por ej todas las especies de Helechos
– Traqueofitas Angiospermas, sus óvulos están cubiertos o protegidos por el Ovario de la Flor. Las Angiospermas se subdividen en 2 grupos:
1- Monocotiledóneas, en donde sus Semillas están constituídas por un solo Cotiledón, por ej el Maíz
2- Dicotiledóneas, en donde sus Semillas están constituídas por 2 Cotiledones, por ej el Poroto
– Traqueofitas Gimnospermas, en donde los Óvulos están libres o Desnudos(No protegidos por el Ovario de la Flor), por ejemplo todas las especies de Pinos, Abetos, Alerces, Araucarias, Ginkp Biloba.
Caracteristicas:
*se les llama plantas vasculares ya que poseen vasos conductores.
*formado por raiz, tallo, hoja. Su nutricion es autotrofa, producto del proceso fotosintetico.
*desarrolan 3 tipos de tejidos: conductores, mecanicos, tegumentos.
*se pueden producir por semillas. Se clasifican en: *Psilopsila*lycopsida*sphenopsida.
Las plantas vasculares, también conocidas como traqueofitas, se caracterizan por tener, salvo muy pocas excepciones, un plan estructural externo muy homogéneo, donde el cuerpo o cormo está constituido por el vástago, diferenciado en tallo y hojas, y la raíz. Desde el punto de vista anatómico presentan en común: cutícula para limitar la pérdida de agua, estomas para efectuar el intercambio gaseoso, tejidos especiales para proveer soporte, tejidos para la conducción de agua y nutrientes. El alternante dominante en el ciclo de vida es el esporofito. El tallo o eje se considera el órgano más primitivo de las plantas vasculares, puesto que las más simples están formadas sólo por ejes. Durante la evolución, las partes subterráneas de estos ejes se transformaron en raíz y algunos ejes aéreos sufrieron modificaciones que los convirtieron en los órganos aplanados que son las hojas. Como característica particular presentan xilema, tejido conductor de agua que aparece por primera vez en este grupo. Son caracteres de interés en la separación del grupo: los tipos de hojas y los patrones o modelos (estelas) en que se disponen los tejidos conductores primarios (floema y xilema) en la parte media del tallo o cilindro central.
Se diferencian dos tipos de hojas atendiendo a su evolución filogenética y complejidad alcanzada: micrófilas y macrófilas. La micrófila es el tipo de hoja más simple y su vascularización no está asociada a una interrupción en la continuidad del tejido vascular del tallo, ya que sólo una pequeña porción periférica de éste se desvía para constituir la traza foliar. La macrófila es el tipo más complejo de hoja y su vascularización está asociada a la interrupción del cilindro vascular del tallo, que en el punto donde se inserta la hoja al tallo se desvía completamente para formar la traza foliar, dejando una zona libre de tejido vascular que se rellena con parénquima y que se conoce como laguna foliar Presentan micrófilas los licopodios y equisetos, mientras que las pteridofitas y espermatofitas (gimnospermas y angiospermas) tienen macrófilas. La parte central del tallo y de la raíz en crecimiento primario se conoce como estela o cilindro central y se presenta con diversas formas, modelos o patrones, características de grupos taxonómicos. Está formada por el sistema vascular (floema y xilema) y los tejidos asociados con él: periciclo, radios medulares y médula. La estela es un concepto filogenético y permite establecer relaciones de homología y evolución entre los diferentes grupos de plantas. Las estelas no meduladas aparecen en los grupos más primitivos y están asociadas a micrófilas, mientras que las estelas meduladas aparecen en grupos más evolucionados y están asociadas a macrófilas. En las plantas vasculares la generación dominante en el ciclo de vida es el esporofito, morfológicamente más conspicuo y siempre fisiológicamente independiente, el cual se origina por crecimiento del embrión. Un embrión completamente desarrollado de una planta vascular está organizado en: suspensor, pie, ápice de la raíz, eje embrional, ápice del vástago, y hojas embrionales. En el embrión adulto se pueden distinguir dos zonas de crecimiento o meristemas apicales, que son: el meristema ápical del vástago (tallo y hojas), y el de la raíz (subapical), que normalmente persisten durante todo el crecimiento de la planta. Estos meristemas son responsables de la conformación final del esporofito adulto al formar un sistema de tallos y hojas, y un sistema radical. Al alcanzar el estado fértil, el esporofito comienza a producir esporas (meiosporas) en estructuras localizadas que son los esporangios. Las esporas pueden ser de dos tipos: 1.- morfológicamente iguales, y en ese caso las plantas que las presentan se denominan isospóricas u homospóricas, o 2.- morfológicamente diferentes, y las plantas serán entonces heterospóricas. Las esporas pequeñas se llaman microsporas (formadoras de gametofitos masculinos en la mayor parte de los casos), y las grandes, macrosporas (megasporas) (formadoras de gametofitos femeninos en la mayor parte de los casos); los esporangios donde se producen se denominan microsporangio y macrosporangio (megasporangio) respectivamente. Los esporangios difieren ampliamente en posición, tamaño, forma y ontogenia. En cuanto a la posición, se pueden encontrar sobre tallos o asociados a esporófilas.
En algunas plantas vasculares inferiores, las esporófilas son semejantes a las hojas estériles o frondes. En otras plantas vasculares inferiores y en las gimnospermas, las esporófilas se reúnen en el extremo de vástagos formando conos o flores estrobiliformes. En las angiospermas (el grupo más evolucionado) es donde alcanzan mayor complejidad y peculiaridad, agrupándose en las flores típicas (flores cíclicas), con dos tipos de esporófilas muy especializados: el estambre y el carpelo. Los gametofitos están muy reducidos en este grupo y se describen dos tipos: exospórico y endospórico. Los gametofitos exospóricos son de vida libre y no están encerrados por la pared de la espora; son típicamente bisexuales, es decir, producen o tienen la capacidad de formar gametangios femeninos y masculinos, y los presentan las plantas homospóricas. Los gametofitos endospóricos están encerrados total o parcialmente por la pared de la espora, restringidos a las plantas heterospóricas y son, por lo general, estrictamente unisexuales. En las plantas vasculares inferiores o pteridofitas, el gametofito masculino está muy reducido, consistiendo corrientemente de una célula rotélica y de un anteridio. El gametofito femenino, al contrario, es mucho más grande y consiste de una masa de tejido almacenador de alimentos que llena la cavidad de la macrospora y queda en parte expuesto al romperse su pared. En la zona expuesta se forman los arquegonios. Las plantas vasculares superiores o espermatofitas son siempre heterospóricas y sus gametofitos son endospóricos. En este grupo hay una fuerte tendencia a la disminución del número de células estériles o vegetativas, por lo que los gametofitos son muy pequeños (con pocas excepciones microscópicos). La macrospora queda retenida, produciendo un gametofito femenino que depende del esporofito desde el punto de vista nutritivo. Los arquegonios solamente se encuentran en las gimnospermas, ya que en angiospermas la reducción del gametofito femenino es tan acentuada que no se forman arquegonios. El microsporangio produce microsporas (granos de polen) que son generalmente liberadas ya germinadas con el gametofito masculino, que es muy reducido (2 o 3 núcleos, sin anteridios). Los granos de polen deben alcanzar la estructura receptora femenina por polinización y sobre ella el gametofito masculino continúa su desarrollo y se prolonga en un tubo polínico, a través del cual se desplazan los gametos masculinos.
En las plantas vasculares con gametofitos de vida libre, las pteridofitas, la sexualidad es referida a los gametofitos, y se presentan dos condiciones: la dioica y la monoica. En las espermatofitas, donde los gametofitos no son plantas de vida libre, están muy reducidos y el femenino queda retenido en la planta esporofítica, la sexualidad hace referencia a la forma en que se distribuyen los microsporangios y macrosporangios en las plantas esporofíticas. Se presentan tres condiciones fundamentales: dioica, monoica y hermafrodita.
LOS HELECHOS:
1. ORIGEN
Los helechos son plantas vivaces, originarias de zonas ecuatoriales y tropicales húmedas (Adiantum, Asplenium, Platicerium…), de regiones tropicales y subtropicales (Nephrolepis, Pteris…) y de regiones donde el clima es de tipo mediterráneo (Blechnum, Cyrtomium, Pellaea…).
2. IMPORTANCIA ECONÓMICA Y DISTRIBUCIÓN GEOGRÁFICA
Debido a la belleza de su follaje, las Pteridofitas ornamentales se han hecho muy populares tanto en arreglos florales (confección de ramos, centros y composiciones florales, ya que sus hojas ofrecen una adaptabilidad a la confección de estos, como elementos centrales o complementarios) así como en jardines y en plantas de interior. La especialización en la producción de jóvenes helechos se generaliza cada vez más, al tratarse de plantas habitualmente resistentes, a condición de no exponerlas excesivamente ni al sol ni a la sombra y que tengan una temperatura constante entre 12 y 18ºC. Hasta mediados del siglo XX, los helechos eran muy populares como plantas de interior, pero debido a la introducción de la calefacción central en las viviendas, esta popularidad disminuyó, pues el aire seco es perjudicial para los helechos. Actualmente la aclimatación ha mejorado mucho y la gente ha vuelto a apreciarlos. Sobre todo los lugares frescos como miradores, cuartos de baño… son muy aptos para los helechos.
3. TAXONOMÍA Y MORFOLOGÍA
Los helechos son plantas sin flores ni semillas, pertenecientes al grupo de las Pteridofitas. Se reproducen mediante esporas, las cuales necesitan la presencia de agua para completar su ciclo biológico. Los helechos son notables por:
Sus hojas (frondes).
Su tallo subterráneo (rizomatoso).
Su reproducción particular.
Sus numerosos géneros y especies.
Una vez que alcanzan el momento de la reproducción en algunos frondes se generan los cuerpos reproductores, denominados soros, donde están los receptáculos de esporas, llamados esporangios. La forma de situarse los soros en los frondes suele ser una característica específica y el conocimiento del estado de madurez de los esporangios es de gran ayuda para su reproducción.
4. LOS HELECHOS MÁS CULTIVADOS
4.1. Adiantum
-Origen: Brasil.
-Especie: Adiantum raddianum.
-Cultivares: "Fragantissima", "Brilliantelse", "Sabina", "Monocolor".
-Características: crecimiento rápido, con pecíolos de frondes negras, brillantes y finas. Las pínulas ("hojas" ) son más o menos triangulares. Se suelen cultivar en invernadero climatizado. Requieren por lo general ambiente húmedo y no soportan excesos de temperatura. Se utilizan tanto como planta de interior como para verde de corte.
4.2. Asplenium
-Orígenes: India-Australia.
-Especie: Asplenium nidus (nido de ave).
-Características: helecho epífito de crecimiento lento, con frondes erectos, ovado-lanceolados, de 150 X 20 cm, estrechándose poco a poco hacia la base, son simples, enteros, de textura algo coriácea, de color verde amarillento brillante, con fuertes nervios negros.
Se cultiva en invernadero climatizado.
Es una magnífica planta de interior, aunque no soporta los ambientes excesivamente secos.
4.3. Nephrolepis Publicidad Curso Especilista en
Jardinería
En este curso se describen los diferentes componentes que componen un jardín, así como las técnicas para ejecutarlo y mantenerlo adecuadamente. Este curso pretende explicar de forma clara y sencilla el diseño, implantación y manejo de un jardín.
-Origen: América tropical (N. exaltata) y Extremo Oriente (N.cordifolia).
-Especies:
Nephrolepis exaltata: con rizoma bien desarrollado, escamoso, sin tubérculos. Frondes de 70-100 x 5-12 cm, con pecíolo moderadamente escamoso. Pinnas 2.5-6 x 0.6-1.4 cm, triangular-oblongas, en ocasiones algo falcadas, con la base truncado-auriculada o auriculada. Cuenta con numerosos cultivares "Teddy Junior", "Boston", "Maasii", "Rooselvetii", "Emina".
Nephrolepis cordifolia: con rizoma bien desarrollado, escamoso, con escamas a veces rojizas o pardas. Frondes de 25-90 x 3-7 cm, con pecíolo algo escamoso. Pinnas de 1-3 x 0.5-0.9 cm, de oblongas a lanceolado-oblongas, rectas o ligeramente falcadas, con la base auricular-cordiforme. Son menos cultivadas que la anterior especie.
-Características: Nephrolepis deriva de las palabras nephros (riñón) y lepis (escama). La "escama reniforme" hace referencia a los indusios en forma de escama sobre las esporas.
Son de crecimiento rápido.
Se cultiva en invernaderos climatizados y muy luminosos.
Es el helecho en maceta más importante en cultivo; además es un excelente planta colgante.
4.4. Platycerium
-Orígenes: Australia, África y América tropical.
-Especies:
Platycerium bifurcatum: (helecho cuerno de alce). Helecho con frondes estériles de 60 x 45 cm, sésiles, erectos, redondeados, enteros y ondulados o lobulados, de textura papirácea. Frondes fértiles de 90 cm de longitud, colgantes, 2-3 bifurcados de textura coriácea, con segmentos acintados, obtusos, pubescentes.
Platycerium superbum: (helecho cuerno de ciervo). Helecho con frondes estériles de 160 x 150 cm, sésiles, cuneados, con el ápice truncado, bifurcados 4 veces o lobulados, de color verde grisáceo. Frondes fértiles de 2 m de longitud, colgantes, cuneados, 5 veces bifurcados, con segmentos a menudo enrollados.
-Características: helecho epífito de frondes características y de crecimiento lento. Algunos de los frondes modificados se adaptan para hacer el soporte de la planta. Se suelen cultivar adheridos a troncos de forma lateral o en pequeñas cavidades a los lados de un eje, lo que resalta su valor ornamental.
Se suelen cultivar en invernadero climatizado.
4.5. Pteris
-Orígenes: Europa meridional (P. cretica) y Extremo Oriente (P. serrulata y P. ensiformis).
-Especies: se trata de un género muy comercial.
Pteris cretica: helecho con rizoma escasamente rastrero. Frondes de 15-80 cm de longitud, con pecíolo más largo que la lámina y con la base truncada. Pinnas basales enteras o aserradas, lineares. Cuenta con numerosos cultivares "Albo-lineata" (rayada de blanco), "Parkeri" (porte más o menos erecto, de color verde oscuro y vigoroso)
Pteris serrulata: parecidas a la especie anterior. Cultivar "Ouvrardii".
Pteris ensiformis: helecho con frondes bipinnadas, de 15-30 cm de longitud, con la pinna terminal de 5-10 cm de longitud, ligeramente compuesta. Pinnas laterales de 4-5 pares, divididas en 2-6 pínnulas obovado-oblongas, dentadas. Cultivar "Evergemiensis" de pínulas finas, ligeras, verdes y blancas.
–Características: de crecimiento rápido con frondes de pínulas alargadas.
Algunas son muy populares para terrarios, composiciones y pequeñas macetas.
5. MULTIPLICACIÓN
-Por semilla: a partir de esporas maduras (negras), colocadas en bandejas sobre turba morena encalada con pH alrededor de 5.5. Se recomienda distribuir de 1 a 2 g de esporas/m2. La temperatura deberá estar en torno a 22-25ºC. Se debe recubrir con una lámina de vidrio o con un pequeño túnel de plástico y sombrear.
La irradiación de la semilla y de las plantas jóvenes en invierno pueden permitir una producción más regular. Aplicar de 1.000 a 2.000 lux, es decir, de 80 a 100 W/m2 (por ejemplo, con tubos fluorescentes). Iluminar a la caída del día (duración del día + iluminación= 16 horas).
Se deberá asegurar un grado de higrometría suficiente: 80% o más.
-Por separación de estolones: se emplea en el género Nephrolepis, pues éstos emiten largos y finos estolones que poseen yemas terminales que al entrar en contacto con el suelo, generan hijuelos. Estos pueden ser separados y empleados para el cultivo en maceta.
Los estolones son extraídos de las plantas madre cultivadas en invernadero climatizado (a 25ºC de mínima) sobre bandejas, en un sustrato a base de turba y mantillo de 30 cm de espesor. El pH de este sustrato lo podemos corregir añadiendo dolomita o hidróxido cálcico. De este modo se puede producir estolones todo el año.
-Por división de plantas: practicada en los géneros Nephrolepis y Adiantum. No es considerado un método industrial.
-Cultivo in vitro: la potencialidad de desarrollo de explantes de primordios foliares y de hojas, en condiciones in vitro, ha sido estudiada para algunas especies de helechos.
Para el cultivar de Nephrolepis "Teddy Junior"; la micropropagación in vitro permite evitar el cultivo de pies-madres sobre superficies importantes para la producción de estolones.
Así mismo se ha utilizado de forma parcial en la germinación de esporas y fase gametofítica de algunas especies propagadas por esporas.
Las plantas madre se seleccionan entre las mejores y más jóvenes. Su explotación no debe prolongarse por más de dos años, al cabo de los cuales se debe renovar con plantas nuevas que pueden proceder del cultivo in vitro.
-Por esporas: este método requiere una instalación muy cuidada; suele llevarse a cabo por establecimientos especializados que venden plántulas de uno o dos repicados.
Los helechos, tal como los conocemos, con raíces, tallos y frondes, son la fase esporofítica.
En las frondes maduras se desarrollan los soros, conteniendo los esporangios donde están las esporas, dando lugar a los prótalos o gametofitos. Estos prótalos desarrollan dos tipos de órganos especializados: los anteridios, que producen los anterozoides y los arquegonios, que contienen el óvulo. El anterozoide, en presencia de agua, fecunda el óvulo, y se produce el nuevo esporofito, que suele surgir desde el prótalo, dando lugar al nuevo helecho.
6. REQUERIMIENTOS EDAFOCLIMÁTICOS
-Temperatura: de 19 a 20ºC en el sustrato. Aunque la temperatura óptima de crecimiento de Asplenium nidus es 16-25ºC, aunque tolera bien temperaturas de 12ºC.
-Iluminación: luz complementaria (ver siembra): 2.000 lux. Asplenium nidus requiere entre 15.000 y 30.000 lux.
Nephrolepis exaltata puede ser cultivado en niveles altos de luz (35.000 a 45.000 lux) si se garantiza un suministro adecuado de agua y nutrientes.
–Humedad relativa: higrometría elevada.
-Suelo: los sustratos deben ser porosos (el porcentaje de porosidad libre óptimo para el cultivo de helechos debe ser mayor del 20%), ligeros y humíferos. Conviene utilizar sustratos con elevada capacidad de almacenamiento de agua, pues las plantas evaporan importantes cantidades.
Se recomienda que el sustrato para helechos esté neutralizado, pudiendo oscilar su pH 4,2/5-5,2/6.
Los valores más adecuados de pH en tres casos particulares son:
Adiantum: 5/5,2-5,7/6
Platycerium: 5,5-6,5
Pteris: 4,7-5,2
Ejemplo de sustrato:
Mantillo (50%).
Tierra de brezo (50%).
O bien:
Turba rubia: 70%
Turbia parda: 20%.
Arena: 10%.
-CO2: raramente aplicado. Es tóxico para Asplenium a 700 ppm.
7. PARTICULARIDADES DEL CULTIVO
-Plantación
Las plantas estarán separadas unas de otras unos 35-45 cm, y su follaje nunca debe cubrir o dar sombra a las nuevas plántulas, por tanto, regularmente se podarán las hojas que molesten al perfecto desarrollo de las plantas. Por supuesto, este marco variará según el tamaño de la planta y la maceta utilizada.
-Repicado
El primer repicado se realiza en estado de prótalo. Se realiza en bandejas, de cuatro a seis meses después de la siembra.
El segundo repicado tiene lugar en estado de 2-3 hojas. Se realiza en bandejas o en bandejas múltiples, de dos a cuatro meses después del primer repicado. En este estadio es cuando comienza la producción a partir de vitroplantas.
–Trasplante
En macetas, con láminas de polyestireno, o bien sobre bandejas de invernadero, seis u ocho meses después. Se colocan de una a tres plantas por maceta (o alveolo) del 10-12.
-Abonado
Se realizarán aportes de abono soluble (equilibrio 1-0.5-1) cada ocho días a una dosis de 0.5 g/l.
La relación nitrógeno/potasio debe ser cuidada, pues un exceso de nitrógeno puede originar malformaciones (bordes lobulados) una aportación de 100 ppm, utilizando un equilibrio 3:2:3 puede ser aconsejable; además se pueden intercalar abonos foliares, sobre todo en invierno.
Nutrientes en kg/ha en helechos

-Riego
Cualquier sistema de riego localizado es aconsejable.
Nephrolepis exaltata soporta mal el riego directo sobre los frondes.
Asplenium nidus requiere mucha cantidad de agua, especialmente durante el periodo estival, pero hay que tener en cuenta que la humedad no debe ser excesiva en la base de la planta.
8. PLAGAS Y ENFERMEDADES
8.1. Plagas
-Cochinillas
Es la plaga más habitual de los helechos.
Los frondes aparecen cloróticos, deformados y pegajosos debido a la melaza que excretan las propias cochinillas.
Control:
-Se realizarán tratamientos a base de Oleoparation.
-Los compuestos organocúpricos pueden causar fitotoxicidad.
-Nemátodo foliar (Helenchoides fragariae)
Provocan graves daños en las hojas, se reconoce por el color pardo rojizo de la base de los frondes.
Control:
-Tratamientos con Aldicarb, Oxamilo o Fenamifos.
-Destruir las plantas que evidencian síntomas de severo ataque.
-Se pueden incorporar insecticidas al sustrato en el momento de los transplantes de forma granulada.
8.2. Enfermedades
–Bacterias (Pseudomonas)
Causan necrosis en las hojas más jóvenes.
Sobre Asplenium nidus presenta manchas acuosas translúcidas, localizadas en el margen de las hojas más tiernas.
Control:
-Eliminar el riego aéreo.
-Suprimir las plantas infectadas.
-Tener un buen control del riego en invierno.
-Botrytis
El exceso de humedad o el riego directo puede ocasionar ataques de Botrytis sobre los frondes de Nephrolepis exaltata.
Control
-Conviene tratar preventivamente con Benomilo.
-Pythium
Ataca sobre todo las raíces de las plantas jóvenes.
Control
-Se puede prevenir con Clortalonil.
-El uso de Propamocarb, inhibe temporalmente el crecimiento de los frondes de Nephrolepis exaltata, por tanto no se aconseja su uso.
–Virus
-Fern virus: es transmitido por áfidos infectando al género Pteris, siendo el responsable de graves deformaciones foliares.
-Impatiens necrotic spot tospovirus: infecta al género Asplenium, produciendo manchas necróticas foliares en forma de anillos.
Para su comercialización, si el transporte es por avión o contenedor, se debe tener en cuenta el tiempo que tardará desde su salida del invernadero hasta su llegada a destino y realizar un correcto empaquetado, teniendo en cuenta que un exceso de humedad y falta de ventilación pueden hacer que a su llegada estén convertidos en productos de desecho.
Se aconseja comercializar Nephrolepis en fundas, ya que se protege las plantas contra daños y se limita la evaporación. Además se aprovecha mejor el espacio durante el transporte. Pero, al almacenar las plantas, es mejor quitar las fundas temporalmente.
Características particulares de los Helechos:
*existen unas 10.000 especies.
*son plantas herbaceas y risomatoza.
*viven en ambientes regionales calidos y humedos, especialmente en selvas tropicales.
*tienen verdaderas raices, tallos, hojas.
*las hojas presentan venas o lenquinas epidermis protectoras y estomas.
*las esporas se producen en los esporaneos que se encuentran agrupados en una estructura llamada Soros.
*en el tallo tienen una capa llamada epidermis q le sirve de proteccion, tejidos de soporte y vasculares.
Caracteristicas Gimnosperma: *plantas con semillas.
Reciben el nombre de coníferas *no tienen capa protectora, ni fruto que la proteja, se les llama tambien plantas de semillas desnudas. Estas semillas son esenciales para la reproduccion por que tienen un embrion, un tejido que permite el crecimiento del embrion debido a las reservas alimenticias y una cubierta que la proteje. *sus hojas son consideradas fructiferas.
Caracteristicas Angiosperma:
Son plantas con flores y frutos. *tienen ovulos que se convierten en semillas para producir la fecundacion y la formacion del fruto. *en la misma flor se puede encontrar el aparato reproductor femenino y masculino de esas plantas.
Ciclo vital del helecho: En la parte inferior del prótalo se diferencian los anteridios y arquegonios. Los primeros liberan un gran número de gametos masculinos flagelados que moviéndose el agua se trasladan hacia los arquegonios atraídos por sustancias hormonales segregadas por los gametangios femeninos. Una vez alcanzado el arquegonio penetran en él a través de cuello y fecundan la oosfera que está situada en la parte basal ensanchada del arquegonio. Esto da origen a una pequeña lámina muy simple, formada por celulas haploides, que constituye el prótalo. Importancia de los Helechos: su importancia es limitada. Algunos sin embargo, encuentran un cierto uso en la industria farmacéutica por los principios vermífugos que contienen. En algunos países, ciertas especies de helechos proveen materiales comestibles ricos en almidón. Gran importancia tienen muchos helechos cultivados como plantas ornamentales.
Adaptacion: es la formación de hojas compuestas llamadas ESPOROFITO. Además en el envés de estas hay una serie de elementos circulares ( SOROS ) y dentro se encuentran los ESPORANGIOS, portadores de las células reproductoras asexuales llamadas ESPORAS. Necesitan además de un Tallo llamado RIZOMA que crece horizontalmente a la superficie del suelo y acumula sustancia de reserva y de él se deprenden las RAÍCES adventicias y fibrosa.
PARTES DE UN HELECHO:

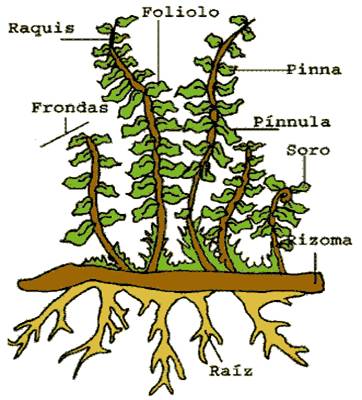
TIPOS DE FONDAS:

Los helechos poseen vasos conductores, por lo tanto, son capaces de transportar las sustancias nutritivas por toda la planta. En una planta de helecho se pueden distinguir unas raíces pequeñas que salen de un tallo subterráneo llamado rizoma, el cual se encuentra recubierto por unas pequeñas escamas. Por el otro lado del rizoma salen las frondas u hojas de helecho.
REPRODUCCIÓN: Cuando los esporangios han madurado, se abren y las esporas son liberadas, caen al suelo y si las condiciones son buenas, es decir, hay humedad y calor, germinan y dan lugar a una pequeña hoja en forma de corazón llamada prótalo. Si decides buscar un prótalo, te costará mucho trabajo encontrarlo ya que son bastante pequeños y no duran mucho tiempo. Son importantes para los helechos ya que en ellos se encuentran los órganos reproductores femeninos y masculinos.

Hábitat: Los helechos son plantas (por lo tanto recuerda que son autótrofas y que elaboran su propio alimento) a las que les gustan los lugares muy húmedos y con sombra. Es común encontrarlos a orillas de ríos, riachuelos y acequias o en quebradas profundas.
Espermatófitas:
La adquisición de semillas representó la cúspide de la adaptación al medio terrestre en el reino vegetal. En las plantas con semilla ya no solo predominó el esporofito, sino que el gametofito se ha reducido hasta llegar a ser una estructura dependiente encerrada en el arquegonio del esporofito.. Por otro lado los espermatozoides flagelados de las plantas inferiores fueron sustituidos por el proceso de polinización. Fue la polinización lo que permitió que las plantas se independizaran del agua como vehículo de fecundación. Asimismo, el cigoto y el embrión en desarrollo (que se convertirá en el nuevo esporofito) dejaron de depender del agua, pues la semilla aloja al embrión dentro de una dura cubierta y lo protege contra otros mecanismos.
Las espermatófitas modernas se dividen en: Gimnospermas y Angiospermas.
Gimnospermas
Las gimnospermas (del latín, gymn-, "desnuda"; del griego, sperma, "semilla") (foto 6 y 7), nombre que reciben las plantas vasculares que forman semillas pero carecen de flores. Comprenden varios grupos: cicadofitos, ginkgos, coníferas (foto 8) y gnetofitos. Las gimnospermas son plantas leñosas de porte arbustivo, arbóreo o, más raramente, trepador (algunas plantas gnetofitas). Se diferencian del otro filo de plantas con semillas, el formado por las plantas con flores (Angiospermas), en que las semillas no están encerradas en carpelos, sino dispuestas sobre escamas organizadas en conos. Las gimnospermas son las plantas con semillas más antiguas; al parecer, proceden de helechos del devónico. Los cicadofitos conservan los caracteres más primitivos de las actuales plantas con semillas. Las pruebas morfológicas y moleculares sugieren que las gnetofitas comparten un antepasado común con las plantas con flores. Las gimnospermas vivientes están distribuidas por todo el mundo, pero prefieren, en particular las coníferas, las regiones templada y subártica. Cicadofitos y gnetofitos son primordialmente tropicales y subtropicales. Hay unos 70 géneros y 600 especies de gimnospermas vivientes, muchas menos que en muchas familias de plantas con flores.
¿Cómo se reproducen las gimnospermas?
Analicemos el ciclo vital de un pino (figura 1), una de las gimnospermas más conocidas por todos nosotros. Los pinos pertenecen al subgrupo de las coníferas.
En el ciclo vital de estas plantas, el árbol corresponde a la generación esporofítica 2n y los gametofitos n consisten en un número pequeño de células que se originan en el esporofito.
Todas las coníferas producen dos tipos de esporas que se desarrollan en las piñas o conos masculinos y femeninos. Los conos masculinos son más pequeños que los femeninos y se encuentran en las ramas más bajas del árbol. Los conos femeninos están ubicados en las ramas más altas.
Los conos femeninos están formados por unas hojas leñosas que poseen en sus bases células formadoras de esporas haploides, que dan origen al gametofito femenino. Los conos masculinos están formados por estructuras superpuestas en forma de hojas. En la base de cada una de estas hojas se encuentran las células formadoras de esporas haploides. Estas esporas dan origen a gran cantidad de granos de polen o gametofitos masculinos que son transportados a través del viento. Cuando caen en los conos femeninos, se desarrolla en los granos de polen un largo tubo polínico que pone en contacto el núcleo masculino con el femenino. Este proceso puede llegar a durar hasta un año. Una vez realizada la fecundación de los núcleos, se forma una cigota 2n que se divide mitóticamente y origina el embrión del esporofito. El gametofito femenino rodea el embrión y pasa a formar parte del material nutritivo en la semilla madura. Tanto el embrión como el material nutritivo quedan rodeados por el tegumento que protege a la semilla. Cuando el cono femenino se abre, deja en libertad a las semillas que poseen un ala de tegumento que facilita su dispersión a través del viento.
Si las semillas caen sobre la tierra y en un ambiente adecuado, germinan y originan nuevos esporofitos.
Las gimnospermas presentan varios avances evolutivos sobre los helechos. Por ejemplo:
a) la fecundación no depende del agua;
b) la formación de un tubo polínico que guía el encuentro de los núcleos femenino y masculino, facilitándose así las posibilidades de una fecundación exitosa;
c) al permanecer el gametofito femenino en el esporofito, la cigota y el embrión poseen mayor protección en sus primeras etapas de desarrollo;
d) el embrión se encuentra protegido de la deshidratación y de posibles daños mecánicos por la semilla. De esta manera, el embrión puede permanecer en estado de latencia hasta que las condiciones ambientales sean las adecuadas para la germinación de la semilla y alimentarse de los nutrientes presentes en esta estructura. Todas estas características hacen que las gimnospermas sean las primeras plantas cuya reproducción está totalmente adaptada a la vida terrestre.
Angiospermas
Las angiospermas (del latín angi-, encerrada, y del griego sperma, semilla), nombre común de la división o filo que contiene las plantas con flor (foto 9), que constituyen la forma de vida vegetal dominante. Los miembros de esta división son la fuente de la mayor parte de los alimentos en que el ser humano y otros mamíferos basan su subsistencia, así como de muchas materias primas y productos naturales. Pertenecen a este grupo casi todas las plantas arbustivas y herbáceas, la mayor parte de los árboles, salvo pinos y otras coníferas, y plantas más especializadas, como suculentas, epifitas y acuáticas. Aunque se conocen cerca de 230.000 especies, hay muchas todavía ignoradas. Las plantas de flor han ocupado casi todos los nichos ecológicos y dominan la mayor parte de los paisajes naturales. Aproximadamente las dos terceras partes de todas las especies son propias de los trópicos, pero las actividades humanas las están exterminando a gran velocidad. Sólo un millar de especies tienen importancia económica digna de consideración, y el grueso de la alimentación mundial procede de sólo quince especies. Si se investigaran como es debido, podrían utilizarse varios cientos más de especies. Se presentan dos grandes grupos principales de plantas con flores: Monocotiledóneas y Dicotiledóneas.
Descripción de las angiospermas:
El elemento más característico de las angiospermas es la flor, cuya función es asegurar la reproducción de la planta mediante la formación de semillas. Las flores son hojas muy modificadas, formadas por cuatro partes fundamentales dispuestos en series independientes, o verticilos, que crecen en el ápice de tallos especializados. El verticilo externo es el cáliz, o conjunto de los sépalos, que son hojas modificadas, o brácteas, casi siempre de color verde. Viene a continuación la corola, formada por los pétalos; éstos también son hojas modificadas, pero normalmente de textura más fina y color más vivo. El tercer verticilo corresponde a los estambres, piezas formadoras de polen, que en conjunto constituyen el androceo o porción masculina de la flor. El verticilo más interno es el gineceo, formado por los carpelos, las estructuras femeninas encargadas de formar las semillas; los carpelos suelen fundirse en una estructura llamada estilo, terminando en un extremo libre llamado estigma.
Las semillas de las angiospermas se desarrollan dentro de un ovario, la parte del carpelo que rodea y protege los óvulos reproductores. La semilla se forma a partir del óvulo después de la polinización y la fecundación (fig. 3). Ni óvulos ni semillas son elementos exclusivos de las angiospermas; las plantas de semilla desnuda como mencionamos anteriormente (Gimnospermas), tienen óvulos que permanecen expuestos en la superficie de las hojas escamiformes especializadas de las piñas o conos. Lo exclusivo de las angiospermas es el desarrollo de la semilla a partir de un óvulo encerrado en un ovario que, según crece la semilla fecundada, se agranda hasta convertirse en fruto.
Evolución de las angiospermas:
La exclusiva flor de las angiospermas probablemente evolucionó a partir de una gimnosperma ahora extinguida con piñas, polinizadas por insectos, que combinaban partes reproductivas masculinas y femeninas. Casi todas las gimnospermas vivientes tienen polinización anemófila (por el viento), pero algunos grupos fósiles y supervivientes aportan indicios de polinización por insectos (entomófila). Aunque los insectos favorecen la polinización, también devoran los óvulos, y se piensa que el desarrollo del carpelo que encierra el óvulo es una adaptación para proteger a éste y a la semilla en desarrollo de la predación de los insectos. El carpelo también protege de otras influencias nocivas del medio, como la desecación, y ha permitido reducir el tamaño del óvulo, perfeccionar el proceso de la polinización y desarrollar otras piezas de la flor, todo lo cual aumenta las probabilidades de reproducirse con buenos resultados.
Ciclo de vida y estructuras reproductivas de espermatofitas:
Las espermatofitas pueden definirse como traqueofitas con las siguientes características:
Como en todas las traqueofitas, hay dos generaciones multicelulares alternadas, denominados gametofito y esporofito, siendo el gametofito taloide, y el esporofito organizado en tejidos y órganos. El esporofito es un "cormo" (con sistema vascular, raíz y vástago). Como en todas las eufilofitas, el vástago del esporofito crece gracias a su meristema apical, y se ramifica como un tallo principal con ramas laterales, y además tiene hojas ("eufilos").
En las espermatofitas la alternancia de generaciones gametofítica y esporofítica se da en forma enmascarada, ya que el gametofito se desarrolla por completo dentro de las estructuras del esporófito (incluso en los grupos más primitivos todavía pueden observarse anteridios y arquegonios);.
Poseen un ciclo de vida heterospórico, con gametofito de desarrollo endospórico (el esporofito da esporas femeninas dentro de las cuales se desarrolla el gametofito femenino, y esporas masculinas dentro de las cuales se desarrolla el gametofito masculino)Los esporangios nacen siempre en las hojas (las hojas fértiles son llamadas "esporofilos"). En las espermatofitas, las hojas fértiles, portadoras de esporangios, siempre están sobre una rama de crecimiento limitado ("braquiblasto"). Por lo tanto en el cormo aparece una nueva estructura, característica de las espermatofitas: la flor (aquí definida como una rama de crecimiento limitado portadora de hojas fértiles). Morfológicamente en la "flor" puede diferenciarse un eje que se origina en el vástago y que presenta una o 2 brácteas, que remata en un receptáculo donde se ubican los megasporofilos que darán las megasporas, y los microsporofilos o estambres que darán las microsporas. Los megasporofilos y los microsporofilos, en conjunto son los antofilos, por eso a este taxón también se lo conoce como Antophyta (pero no es recomendable utilizar ese nombre, porque algunos autores consideran como "antofitas" sólo a las angiospermas y los gnetales, que antes se creía que tenían un antecesor común). En la "flor" veremos el desarrollo "enmascarado" del gametofito:
en la flor las hojas fértiles portadoras de megaesporangios (los "megaesporofilos") contendrán el megaesporangio dentro del cual se produce la meiosis. 3 de los productos de la meiosis degeneran y el sobreviviente se desarrolla en la única espora femenina o "megaspora" del megasporangio, que a diferencia de lo que pasa en las demás plantas vasculares, nunca abandona al megasporangio, es una característica distintiva de las espermatofitas. La megaspora produce en su interior el gametofito femenino, que en su estado adulto puede ser de pocas (6) o muchas células, que tampoco abandona nunca a la megaspora ("gametofito endospórico", fenómeno común en plantas heterospóricas). El gametofito femenino a su vez produce en su interior una o varias gametas femeninas o "huevos" (como en todas las embriofitas, la gameta femenina nunca abandona al gametofito femenino). A esta nueva estructura (megasporangio + megaspora + gametofito + gameta) se la llama óvulo.
La cubierta del megasporangio por mitosis va formando las cubiertas protectoras del futuro embrión (llamadas tegumentos, 1 en gimnospermas y 2 en angiospermas), puede haber una parte del megasporangio que se transforme en una cubierta de tejido de reserva (nucela). Como en espermatofitas la megaspora nunca abandona al megasporangio, el megasporangio de espermatofitas posee una abertura relativamente especializada por la selección natural para recibir el grano de polen y lograr la fecundación. Esta abertura del megaesporangio se llama "micropila". En gimnospermas el óvulo y su micropila están expuestos, en angiospermas están cubiertos por el carpelo (hoja fértil portadora de esporangios). El gametofito femenino produce las gametas femeninas cerca de la abertura de la micropila. en la flor las hojas fértiles portadoras de microesporangios (los "microesporofilos") se llamarán estambres y desarrollarán el microsporangio (que en los estambres de las espermatofitas se llama "saco polínico") que dará las microsporas, cada una contendrá un solo gametofito masculino (el gametofito masculino maduro cubierto por la pared de la microspora es el "grano de polen"). Los granos de polen serán liberados al exterior por "polinación" (apertura del saco polínico o microsporangio).
Se da el nuevo fenómeno de polinización que es el transporte del polen hasta las estructuras de la flor femenina preparadas para recibirlo, mediante agentes externos. En gimnospermas el polen se transporta directamente a la micropila, en angiospermas se transporta al estigma del carpelo. Los agentes externos son principalmente el viento (fenómeno de anemofilia, común en gimnospermas) o los animales (fenómeno de zoofilia, común en angiospermas).
Cuando la polinización es exitosa, el gametofito masculino crece, atraviesa la pared del grano de polen y emite un tubo polínico o haustorial, que emitirá los anterozoides poliflagelados –carácter primitivo- o los núcleos espermáticos -carácter avanzado- en el óvulo produciéndose la fecundación.
Luego de la fecundación el gametofito femenino por mitosis va formando un tejido nutricio de reserva (haploide en gimnospermas, llamado protalo o endosperma primario, triploide en angiospermas por fecundación con un núcleo espermático del polen, llamado endosperma a secas)
Como en todas las embriofitas, el cigoto por mitosis se transforma en un embrión, que creció nutrido por el gametofito femenino a través de una placenta. En las espermatofitas, el embrión al madurar entra en estado de latencia rodeado de las estructuras del gametofito -protalo o endosperma- y esporangio femeninos -nucela o no, 1 o 2 tegumentos- formando la semilla. el embrión es bipolar con meristema apical y meristema radical.la semilla madura es liberada convirtiéndose en la unidad de dispersión (a diferencia de la espora de pteridophytas) en condiciones adecuadas la semilla germina (despierta de la latencia), la primera fase de la germinación es la absorción de agua y luego se sigue desarrollando el embrión por mitosis, nutrido al principio de los tejidos de reserva de la semilla y luego de sus propios productos de la fotosíntesis, dando el nuevo esporofito adulto.
Evolución de espermatofitas:
Como se puede observar de sus hermanos vivientes los helechos, las espermatofitas descienden de un ancestro caracterizado por la homosporía (un solo tipo de espora, gametofitos siempre bisexuales). Un paso crítico en el desarrollo de la semilla fue la evolución de la heterosporía: la producción de dos tipos de esporas, las megasporas y las microsporas, que darán los gametofitos femeninos y masculinos respectivamente.
La heterosporía se originó muchas veces en forma independiente en linajes no emparentados de plantas vasculares (hay ejemplos en lycophytas, en equisetopsidas y en polypodiopsidas), en muchos de esos casos el desarrollo de la heterosporía fue seguido de una reducción del número de megasporas funcionales. En la línea que condujo a las plantas con semilla, en el esporofito adulto que desarrolla sus megasporangios, se produce la meiosis de sólo una célula por megasporangio, y por aborto de 3 de los productos de la meiosis, el número de megasporas funcionales es reducido a sólo un megaspora funcional por megasporangio. Esa única megaspora funcional, en el linaje de las espermatofitas fue retenida dentro del megasporangio, desarrollando su gametofito femenino y gameta femenina por completo dentro del megasporangio de la generación esporofítica anterior. Finalmente el megasporangio desarrolló los tegumentos dejando abierto el pequeño agujero llamado micropila.
Nuestro conocimiento de los orígenes de la semilla se basa principalmente en fósiles bien preservados del Devónico tardío y Carbonífero temprano, que fueron llamados "progimnospermas" o "helechos con semilla". Recordemos que la diferenciación entre un tallo principal y ramas laterales ya había evolucionado en el linaje de las eufilofitas. Lo primero que aparece en el linaje que derivó en las espermatofitas, fue encontrado en el Devónico tardío, y es la aparición de troncos muy grandes, con madera bastante similar en su estructura a la de las modernas coníferas. Estos troncos estaban conectados a enormes sistemas de ramas que portaban muchas hojas pequeñas. Archaeopteris, como ahora es llamada, fue descubierta como heterospórica, pero aún sin formar semillas.
La reconstrucción más exacta y la ubicación en el árbol filogenético de Archaeopteris y otras "progimnospermas" como Aneurophyton (Beck 1981, 1988, Beck y Wight 1988), fue fundamental para que los científicos puedan establecer tanto el origen de la heterosporía como el de la producción de madera, y llegar a la conclusión de que los dos eran anteriores a la evolución de la semilla. Por lo tanto es incorrecto afirmar que esos dos fósiles pertenecen a un clado que se llame espermatofitas ("plantas con semilla"), si bien son ancestros de las plantas con semilla actuales. Por lo tanto las espermatofitas y esos dos fósiles pertenecen a un grupo monofilético mayor, que fue llamado "Lignophyta" (Doyle y Donogue 1990), en referencia a que ya producían madera.
Análisis cuidadosos (por ejemplo el de Serbet y Rothwell 1992) han revelado que las primeras semillas estaban situadas en "cúpulas" ("cupules" en inglés) y cada semilla era cubierta por una excrecencia ("outgrowth") de la pared del esporangio que formaba una cámara especializada para recibir el polen, o "cámara polínica" ("pollen chamber" en inglés). Esta estructura probablemente se ayudaba de una secreción de una gota pegajosa ("gota de polinización" o "pollination droplet") para capturar los granos de polen.
Los tejidos del tegumento externo probablemente derivan de una serie de esporangios estériles, que inicialmente tuvieron la forma de una serie de lóbulos en el ápice de la semilla, más que la forma de una micropila diferenciada
Sistemática de espermatofitas
Diversidad de espermatofitas: grupos vivientes
Hoy en día hay 5 linajes vivientes de espermatofitas: las cícadas, los ginkgos, las coníferas, las gnetofitas y las plantas con flores. Los primeros 4 grupos suelen ser agrupados en el grupo Gymnospermae, en referencia a que poseen las semillas "desnudas" o no totalmente cubiertas por el carpelo, en oposición a las angiospermas o plantas con flores, cuyo carpelo cubre completamente a la semilla. Hábito de una cícada.
Cícadas. Fueron las más abundantes y diversas durante el Mesozoico. Hoy quedan alrededor de 130 especies. Las cícadas generalmente poseen un tronco bajo y ancho, con xilema secundario limitado, y hojas compuestas grandes parecidas a las de los helechos o las palmeras. Son dioicos, eso quiere decir que algunos esporofitos sólo portan óvulos y luego semillas, y otros esporofitos sólo portan estróbilos masculinos productores de polen. Los dos tipos de estróbilo son típicamente muy grandes, y en algunos casos de coloración brillante. Asimismo las semillas suelen ser grandes y usualmente tienen un tegumento externo carnoso y coloreado, presumiblemente una adaptación para atraer a los agentes de dispersión vertebrados. Muchas características de las cícadas pueden ser ancestrales, como el polen con tubo haustorial (en lugar de tubo polínico), y el esperma gigante multiflagelado (en lugar de los núcleos espermáticos). Sin embargo, las cícadas poseen características únicas que las alejan de las plantas con semilla ancestrales, que presumiblemente son caracteres derivados, entre los cuales se incluye la pérdida de la ramificación axilar, la presencia de trazas foliares "girdling" ("¿con fajas?"), y la producción de raíces coralloides que albergan cianobacterias fijadoras de nitrógeno. Dentro de las cícadas, los análisis filogenéticos indican que la primera división del grupo fue la que dividió al linaje de Cycas del resto. Por lo tanto Cycas estaría reteniendo algunos caracteres presumiblemente ancestrales, como los que han sido encontrados en parientes fósiles como Taeniopteris, a saber: el tener muchos óvulos nacidos en los márgenes de carpelos (carpelos definidos como hojas fértiles portadoras de óvulos) con mofología de tipo foliar, en lugar de tener dos óvulos por carpelo peltado que los sostiene apuntando hacia el eje del esporofilo (en su "cara adaxial"), que es el carácter que se encuentra en la otra línea. También en Cycas las hojas fértiles portadoras de óvulos no se encuentran agrupadas en estróbilos, como sí lo están en la otra línea.
Rama de Ginkgo biloba.
Ginkgo biloba
Ginkgos. Sólo hay una especie sobreviviente de ginkgos: Ginkgo biloba. Esta especie es muy raramente encontrada en forma silvestre, pero los árboles presentes en los templos de China fueron mantenidos por siglos por los monjes que los habitan, y en los tiempos modernos fue cultivado por el hombre en las veredas de las ciudades. Quizás la característica más distintiva del Ginkgo moderno es la producción de hojas deciduas, con forma de abanico ("flabeladas"), con venación dicotómica. Los ginkgos son bien conocidos en el registro fósil, donde se observa una gran diversidad en la morfología de las hojas. Como las cícadas, los ginkgos son dioicos (esporofitos diferentes portan o bien carpelos o bien estambres -estambres definidos como "hojas fértiles portadoras de sacos polínicos que contienen a los granos de polen"-). Los óvulos nacen en pares sobre ramas axilares que se piensa que son estróbilos reducidos. El tegumento del óvulo se diferencia en una capa externa carnosa (y olorosa) y una capa interna pétrea (dura) que encierra al gametofito femenino. También al igual que en las cícadas, los ginkgos retienen varios caracteres de las espermatofitas ancestrales, como el polen que emite un tubo haustorial (no polínico), y el esperma flagelado capaz de nadar.
Hábito de Pinus.
Coníferas. Hay aproximadamente unas 600 especies de coníferas vivientes. Son árboles o arbustos con madera bien desarrollada y usualmente hojas de tipo aguja. Normalmente las hojas son solitarias, naciendo a lo largo del tallo, pero en los pinos (Pinus) están agrupadas en ramitas pequeñas. Las hojas usualmente tienen adaptaciones adicionales a la sequedad, por ejemplo estomas hundidos. Sin embargo algunas coníferas del Hemisferio Sur (por ejemplo Podocarpus, Agathis) presentan hojas aplanadas y grandes, y en Phyllocladus se observan ramas aplanadas que parecen hojas. Muchas coníferas son monoicas (poseen carpelos y estambres en el mismo esporofito), pero algunos grupos son dioicos: Juniperus, Taxus, y Podocarpus. En los estróbilos masculinos (que en las coníferas se llaman conos masculinos o "pollen cones" en inglés), los estambres (o "microesporofilos") sostienen en su cara abaxial a los esporangios ("microsporangios") que darán gametofitos masculinos ("microgametofitos"). Los granos de polen son los gametofitos masculinos protegidos por una pared originada en el esporofito, y a veces tienen un par de apéndices llenos de aire parecidos a sacos, presumiblemente adaptaciones a la dispersión por el viento, pero estos "sacos aéreos" parecen haberse perdido en muchas líneas. Los óvulos receptivos, a diferencia de los microsporangios, están situados en la cara adaxial de cada carpelo o "escama ovulífera", mirando hacia el eje del cono femenino. La meiosis que dará las gametas, ocurre dentro de cada óvulo, y uno solo de los 4 productos de la meiosis se desarrollará hasta dar el gametofito femenino, siempre dentro del óvulo. El gametofito femenino o "protalo", produce uno o más gametos femeninos o "huevos" en el sector cercano a la micropila. Cuando el grano de polen llega finalmente a la micropila, el gametofito masculino encerrado en él desarrolla un tubo ("tubo polínico") que atraviesa la pared del gametofito femenino. Cuando el tubo polínico termina de crecer, el gametofito masculino emite a través de él dos espermas, que pueden ser células o núcleos celulares ("núcleos espermáticos") según el linaje. Es muy común en coníferas el fenómeno de "poliembronía", con muchos embriones desarrollándose en el mismo gametofito femenino, que puede ser o bien debido a que ocurrieron eventos de fertilización independientes en los que varios huevos fueron fertilizados por varios tubos polínicos distintos, o bien debido a que el embrión único se dividió en una etapa temprana en varios embriones genéticamente idénticos, siendo la última posibilidad más comúnmente encontrada que la primera. En las modernas coníferas, se dice que el estróbilo portador de polen es "simple", mientras que el portador de óvulos es "compuesto". Esto es debido a cómo es interpretada la morfología de los estróbilos: El cono masculino es interpretado como una rama modificada portadora de hojas fértiles o estambres, por lo tanto sería una "flor" única portadora de muchos estambres. En cambio el cono femenino es interpretado como derivado de una rama con hojas, que a su vez porta ramas laterales de crecimiento definido nacidas en la axila de las hojas, cada una de las ramas laterales portadoras de hojas fértiles o carpelos. Esta interpretación es sostenida por el registro fósil, que muestra una serie de pasos en la reducción de las ramas laterales portadoras de carpelos, hasta la aparición de la "escama ovulífera" altamente modificada que vemos en los grupos modernos (Florin 1951, 1954). También se observa que cada escama ovulífera está sostenida por una bráctea ("bráctea tectriz"), que representaría la hoja portadora de la rama lateral, también altamente modificada. En unas pocas coníferas, la bráctea es conspicua, emergiendo de entre las escamas ovulíferas (por ejemplo en Pseudotsuga mensiezii). Sin embargo en muchas coníferas la bráctea es sumamente reducida. En las cupresáceas Taxodium y Cryptomeria, la bráctea está fusionada a la escama ovulífera, y la escama ovulífera aún muestra signos de presentar "hojas" (visibles como pequeños dientes). Los estudios filogenéticos han revelado algunas cuestiones interesantes acerca de la evolución de las coníferas. (por ejemplo Stefanovic et al. 1998). Los datos moleculares muestran una división basal entre las pináceas y un clado que albergaría a todas las demás coníferas. Las pináceas poseen varias características singulares, como óvulos invertidos (con la micropila mirando hacia el eje del cono) y las semillas aladas, alas que se originan en la escama ovulífera durante el desarrollo de la semilla. Dentro del otro clado de las coníferas, los dos grupos más grandes del Hemisferio Sur (Podocarpaceae y Araucariaceae) forman un clado, presumiblemente con la sinapomorfía de poseer un solo óvulo por escama ovulífera. Las cupresáceas están marcadas por muchas características singulares, como la fusión de la escama ovulífera con la báctea tectriz. A su vez, este grupo puede estar emparentado con las taxáceas, que tienen conos femeninos altamente reducidos con una sola semilla terminal, rodeada de un tercer tegumento ("arilo") carnoso y colorido.
Gnetofitas
Gnetofitas. Este grupo contiene sólo unas 80 especies vivientes, que pertenecen a tres linajes bastante diferenciados (Doyle 1996, Friedman 1996, Price 1996). Uno es Ephedra, con alrededor de 40 especies distribuidas en los desiertos de todo el mundo, con hojas muy reducidas escamosas. Otro es Gnetum, con unas 35 especies en bosques tropicales del Viejo y Nuevo Mundo, con hojas con lámina entera muy parecidas a las vistas en la mayoría de las angiospermas. Finalmente, Welwitschia con una sola especie, Welwitschia mirabilis encontrada en el sudoeste de África, produce sólo dos o raramente cuatro hojas funcionales a lo largo de toda su vida, hojas que crecen indefinidamente por meristemas presentes en la base, y se necrosan gradualmente en las puntas.
Hábito de Ephedra.
Hojas y semillas de Gnetum.
Hábito de Welwitschia mirabilis.
Si bien estos 3 clados se ven muy diferentes uno del otro, comparten muchas características inusuales, como las hojas opuestas, múltiples yemas por axila, vasos xilemáticos con aberturas circulares entre células adjuntas, polen compuesto, semillas en estróbilo, y un polen elipsoide ancestral con unas estrías características que corren de punta a punta. Las semillas también tienen dos tegumentos, el interno formando el tubo micropilar que exuda la gota de polinización, el externo derivado de un par de brácteas fusionadas. Los estudios moleculares también consensúan altamente la monofilia de este grupo. Dentro de las gnetofitas, Gnetum y Welwitschia forman un clado bien consensuado. Algunas de las sinapomorfías morfológicas son: hojas con venación reticulada, reducción aún mayor del gametofito masculino, y algunos aspectos de la estructura del gametofito femenino, como el desarrollo tetraspórico, la pérdida de los arquegonios, los núcleos libres funcionando como huevos en lugar de las células. El característico polen estriado encontrado en Ephedra y Welwitschia fue aparentemente perdido en la línea de la que derivó Gnetum, que tiene un polen con granitos con forma de pico, no aperturado. En lo que respecta al registro fósil, es más bien pobre salvo en los granos de polen. Sólo algunos macrofósiles han sido descriptos (Crane 1996). Si bien los granos de polen de las gnetofitas son encontrados desde el Triásico, parece ser que el clado que contiene a los grupos modernos se ha diversificado más significativamente durante el Cretácico medio, al mismo tiempo que las angiospermas. Al igual que las angiospermas, las gnetofitas acortaron su ciclo de vida (y probablemente se volvieron herbáceas) y evolucionaron junto con los insectos para ser polinizadas por ellos, característica aún encontrada en algunos grupos vivientes. En marcado contraste con las angiospermas en cambio, las gnetofitas nunca se volvieron un componente significativo de la flora en paleolatitudes altas y medias, y han sufrido una disminución dramática de su representatividad durante el Cretácico tardío (Crane et al. 1995, Crane 1996).
Flor de angiosperma.
Angiospermas. Con unas 257.000 especies vivientes, las angiospermas son las responsables de la mayor parte de la diversidad en espermatofitas, en embriofitas y en viridofitas. La fuerte evidencia de la monofilia de las angiospermas proviene de los estudios moleculares y de los muchos caracteres morfológicos compartidos por los miembros de este clado. De éstos, algunos de los más obvios, que también son características reproductivas importantes, son: (1) las semillas son producidas dentro de un carpelo con una superficie estigmática que permite la germinación del polen, (2) el gametofito femenino es muy reducido, en la mayoría de las especies son sólo 8 núcleos en 7 células, y (3) la doble fertilización, que llevó a la formación de un tejido nutritivo característico, triploide, llamado endosperma. Otras características son: (4) muchas angiospermas poseen vasos xilemáticos en lugar de traqueidas, carácter derivado dentro del grupo, en los vasos el agua puede fluir sin necesidad de atravesar una membrana, lo que los vuelve muy eficientes en el transporte de fluidos dentro del esporofito pero probablemente también más propensos a recibir daño (en especial por embolias de aire) cuando están sujetos a estrés hídrico. (5) El floema de las angiospermas difiere del de todas las demás plantas en que los elementos del tubo criboso (que son células vivas pero sin núcleo, encargadas del transporte de azúcares) están acompañadas por una o más "células acompañantes", que nacen de la misma célula madre que el elemento criboso.

Las semillas:
La semilla o pepita es cada uno de los cuerpos que forman parte del fruto que da origen a una nueva planta, es la estructura mediante la que realizan la propagación las plantas que por ello se llaman espermatófitas (plantas con semilla). La semilla se produce por la maduración de un óvulo de una gimnosperma o de una angiosperma. Una semilla contiene un embrión del que puede desarrollarse una nueva planta bajo condiciones apropiadas. Pero también contiene una fuente de alimento almacenado y está envuelto en una cubierta protectora.
Estructura: El alimento almacenado comienza como un tejido fino llamado endospermo que es provisto por la planta progenitora y puede ser rico en aceite o almidón y en proteínas. En ciertas especies, el embrión se aloja en el endospermo, que la semilla utilizará para el sexo.
En otros, el endospermo es absorbido por el embrión mientras que el último crece dentro de la semilla en desarrollo, y los cotiledones del embrión se llenan del alimento almacenado. En la madurez, las semillas de estas especies carecen de endospermo. Algunas semillas de plantas comunes que carecen de endospermo son las habas, guisantes, calabazas, girasoles, y rábanos. Las semillas de plantas con endospermo incluyen todas las coníferas, la mayoría de las hierbas y de otras monocotiledóneas, tales como el maíz y el coco.
La envoltura de la semilla se desarrolla a partir de cubiertas, llamadas tegumentos, que originalmente rodean al óvulo. En la semilla esta envoltura madura se puede convertir en una fina cubierta, como en el cacahuete, o en algo más sustancial.
Las semillas de las angiospermas quedan contenidas en estructuras secas o carnosas (o en capas de ambas), llamadas frutos. En español se llama fruta al alimento que representan los frutos carnosos y dulces. En cambio las semillas de las gimnospermas comienzan su desarrollo "desnudas" sobre las brácteas de los conos, aunque en su desarrollo son acompañadas por escamas, que ayudan a su protección o a su dispersión.
Existe también un concepto legal de semillas, en el que se considera como semilla a cualquier parte de la planta cuando su fin es la multiplicación, incluyéndose entonces plantones, vitroplantas, esquejes, etc.

Función de las semillas: A diferencia de los animales, las plantas están limitadas en su habilidad de buscar las condiciones favorables para la vida y el crecimiento. Por consiguiente, han evolucionado de muy diversas formas para propagarse y aumentar la población a través de las semillas. Una semilla debe llegar a la localización adecuada en el momento óptimo de germinación. Estas propiedades que fomentan la producción de la siguiente generación es posible que estén más relacionadas con los frutos que con las mismas semillas, ya que la función típica de la semilla es la de servir de mecanismo retardante, permitiendo suspender el crecimiento si las condiciones no son favorables o dar el tiempo necesario para su dispersión. Cada especie logra su objetivo de una forma diferente: produciendo gran cantidad de semillas, envolviendo las semillas en duras capas que se van ablandando con las lluvias y el frío invernal para germinar.
Producción de semillas: La producción de semillas es un proceso industrial que permite crear semillas de alta calidad, en las mejores condiciones, libres de contaminación, plagas o enfermedades. La producción de semillas es parte de la industria alimentaria (maíz, soya, trigo, frutas entre otros), pero también de productos como el algodón y las flores.
Etapas de producción: La producción de semillas incluye las siguientes etapas: siembra, cosecha, acondicionamiento y secado, análisis y control de calidad, clasificación y selección, lavado y tratamiento, almacenaje, certificación, etiquetado y embalaje, y embarque.
Existen diversos tipos de semilla:
–Girasol: Se extrae de las flores de girasol.
-Marañon: Se extrae de la semilla de marañon.
-Guacalillo: Extraida de los árboles de guacalillo.
-Arroz: Extraído de los árboles de arroz.
-Frijoles: Extraídos de las plantas de frijol.
-Maní: Extraído de los árboles de maní.
-Chan: Es la semilla mas peculiar ya que es extraída de los huevos de los peces como el pez globo, para ello deben estar en tratamiento. No es recomendado ingerirlas ya que pueden contener sustancias venenosas que pueden ocasionar la muerte.
ORIGEN y CARACTERÍSTICAS GENERALES DE LA SEMILLA:
 Página anterior Página anterior | Volver al principio del trabajo | Página siguiente  |