 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
Cinqüenta clones de batata silvestre, tomados ao acaso, da coleção mantida na Embrapa Clima Temperado, foram cultivados no outono e na primavera de 1996. No outono, 30 e, na primavera, 40 clones produziram tubérculos suficientes para a análise de açúcares redutores. Os clones avaliados no outono e/ou na primavera de 1996 estão listados na tabela 1. No outono, foram estudados 24 clones de S. commersonii (19 da subespécie commersonii e cinco da malmeanum) e seis não identificados. Na primavera, foram avaliados outros 11 clones de S. commersonii (sete da subespécie commersonii e quatro da subespécie malmeanum) e um de S. chacoense,, subespécie muelleri. Além dos clones silvestres, três clones cultivados de S. tuberosum,, subespécie tuberosum,, foram utilizados como controle: BR-3, Baronesa e C-1582-25-90, que possuem, respectivamente, alto (1,71%), médio (0,57%) e baixo (0,21%) teor de açúcares redutores (PEREIRA & CAMPOS, 1999).

Os clones silvestres foram plantados sob telado, em sacos plásticos de 8 litros, contendo terra orgânica, com 5kg de adubo NPK por m3. O delineamento experimental foi inteiramente casualizado, com duas repetições por tratamento no de outono e três na primavera. Os experimentos foram plantados em 6 de março e 9 de setembro e colhidos em 27 de julho e 6 de dezembro de 1996, respectivamente, no outono e na primavera.
Os tubérculos produzidos no plantio de outono foram armazenados em condições ambiente após a colheita, por duas semanas, à temperatura mínima variando de 3,2 a 11,7ºC e, à máxima, de 12,2 a 27,4ºC. Os tubérculos produzidos no cultivo de primavera foram armazenados, por doze semanas, à temperatura ambiente, cuja mínima diária variou de 10,8 a 22,0ºC e, a máxima, de 22,6 a 35,6ºC.
Para a análise da concentração de açúcares redutores, os tubérculos foram lavados e deixados à temperatura ambiente, por 24 horas. Após fatiados, foram mergulhados em solução de bissulfito de sódio, por um minuto, e colocados em estufa a 55-65ºC, por 24 horas. Logo em seguida, as fatias desidratadas foram moídas.
As concentrações de açúcares redutores foram determinadas pelo método de Lane Enyon, citado na AOAC (1970), e identificadas pelo método de Somogyi, modificado por NELSON (1944).
As análises estatísticas foram realizadas com o programa SAS (Statistical Analysis System). As análises de variância foram computadas, de acordo com a tabela 1, para estimar as variâncias genéticas ( 2g = QM1 - QM2/r ), ambientais ( 2e = QM2) e herdabilidades (h2).
A herdabilidade foi estimada, no presente trabalho, para determinar a proporção da ação gênica e do ambiente na expressão das características de açúcares redutores, com base na média dos clones, de acordo com a seguinte fórmula (VENCOVSKY & BARRIGA, 1992):
![]()
A concentração média geral de açúcares redutores encontrada nos tubérculos dos clones do cultivo de outono foi o dobro (0,59%) da observada na primavera (0,28%) (Tabela 2). No outono, as concentrações médias de açúcares redutores variaram de 0,16%, para o clone 110, a 2,77%, para o clone 104, sendo que os clones de S. commersonii subespécie commersonii exibiram teores médios de 0,59% e, da subespécie malmeanum, de 0,33%. Vinte por cento dos clones apresentaram níveis de açúcares redutores iguais ou inferiores ao controle para baixa concentração (C-1582-25-90 = 0,26%), sendo, a maioria deles, da subespécie malmeanum (clones 08 e 64 = 0,25%) e de espécies não identificadas (clones: 105 = 0,26%, 110 = 0,16% e 244 = 0,20%). A maioria dos clones exibiu concentração de açúcares redutores intermediária aos controles C-1582-25-90 e Baronesa. Apenas o clone 104 (2,77%) apresentou teor de açúcares redutores mais elevado do que o controle BR-3 (2,20%).

Na primavera, 35% dos clones exibiram teores de açúcares redutores iguais ou inferiores ao do clone C-1582-25-90 (0,21%), predominando, também, clones de S. commersonii malmeanum, os quais exibiram teores médios de 0,22% (Tabela 2). O clone 244 (0,67%) foi o que apresentou a maior concentração de açúcares redutores, com nível semelhante ao controle médio Baronesa (0,63%).
Segundo BEUKEMA & VAN DER ZAAG (1990), são aceitáveis para processamento concentrações de açúcares redutores não superiores a 0,20%, para batata 'chips' e 0,40%, para batata frita na forma de palito, grânulos e flocos. Assim, apenas 6,7% (dois) dos clones estudados estariam dentro dos limites aceitáveis de açúcares redutores para a elaboração de 'chips', no cultivo de outono, e 32,5% (13), no cultivo de primavera. Para batata palito, grânulos e flocos, 46,7% (14) dos clones teriam teores de açúcares redutores aceitáveis, no cultivo de outono, e 90,0% (36), na primavera.
Os valores de açúcares redutores obtidos neste trabalho mostram uma grande variação, principalmente, entre os clones no cultivo de outono e na resposta dos clones aos dois períodos de cultivo, concordando com a assertiva de DALE & MACKAY (1994), quando ressaltam que o conteúdo de açúcares dos tubérculos variam consideravelmente entre cultivares, locais e período de cultivo. Na Região Sul, o cultivo de outono caracteriza-se por fotoperíodos e temperaturas decrescentes, permitindo a exposição dos tubérculos a baixas temperaturas (<10ºC), mesmo ainda antes da colheita, o que propicia a conversão do amido em glicose e provoca um aumento indesejável no conteúdo de açúcares redutores. Esse processo é conhecido por adoçamento de baixa temperatura (AP REES et al., 1981). No presente estudo, a redução da temperatura ocorrida no final do ciclo, no cultivo de outono, deve ter estimulado o acúmulo de açúcares redutores nos tubérculos. Enquanto que, na primavera, as temperaturas ambientais mais elevadas durante a maturação e armazenamento dos tubérculos não favoreceram ao acúmulo de açúcares redutores.
A análise da variância mostrou diferenças significativas no teor de açúcares redutores entre os clones silvestres (Tabela 3), indicando variação para essa característica. Na literatura, não foi encontrado relato de variação genética significativa para açúcares redutores em S. commersonii.
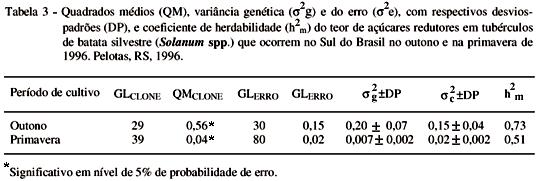
A estimativa da variância genética para açúcares redutores em tubérculos do cultivo de outono foi relativamente grande, sendo de magnitude suficientemente importante para o melhoramento, mesmo que aumentada pela variância do ambiente (Tabela 3). A variância genética no cultivo de primavera foi de baixa magnitude, indicando o pequeno componente genético na população cultivada nesse período. Inclusive, a variância do erro foi superior à variância genética.
A variação quantitativa no número de clones para teor de açúcares redutores no outono (2 clones < 0,20%, 13 clones < 0,40%) foi maior do que na primavera (14 clones < 0,20%, 36 clones < 0,40%), ratificando claramente a maior variância genética para açúcares redutores no outono. Pode-se afirmar, portanto, que é mais apropriado o melhoramento no cultivo de outono, o que é confirmado pelo valor da herdabilidade. Na primavera, não há necessidade de seleção, pois a grande maioria dos clones apresentam as características desejáveis.
A estimativa da herdabilidade para açúcares redutores foi alta no cultivo de outono, e moderada, no período de primavera (Tabela 3). No outono, as temperaturas baixas (< 10º C) induzem a acumulação de açúcares redutores nos tubérculos, amplificando as diferenças entre os clones, facilitando detectá-las. Os valores obtidos são semelhantes aos encontrados por PEREIRA et al. (1994), ao estudar três populações híbridas de Solanum tuberosum, armazenadas a baixas temperaturas (h2 < 0,63).
A constatação de variabilidade genética para açúcares redutores indica a potencialidade das espécies S.commersonii e S. chacoense, como fontes importantes ao melhoramento da S. tuberosum para essa característica. A introgressão dessa característica na espécie cultivada, entretanto, demandará manipulação de ploidia devido à diferença de ploidia existente entre estas espécies silvestres (2n = 2x = 24) e S. tuberosum (2n = 4x = 48).
Com base nos resultados obtidos no presente trabalho, pode-se concluir que as espécies silvestres de Solanum spp. que ocorrem no Sul do Brasil:
a) apresentam variabilidade genética para teor de açúcares redutores, principalmente no outono;
b) possuem herdabilidade moderadamente alta para açúcares redutores no período de outono.
AGRADECIMENTOS
Os autores expressam seus agradecimentos ao grupo de apoio do Programa de Melhoramento de Batata da Embrapa Clima Temperado, pela contribuição na condução dos experimentos no telado, e à laboratorista Eliana Mariete da Luz Silveira, pelo apoio técnico na realização das análises no Laboratório de Fisiologia Vegetal.
AOAC. Official methods of analysis. 11 ed. Washington, D.C.: Association of Official Analytical Chemists, 1970. 1015p.
AP REES, T., DIXON, W.L., POLLOCK, C.J., et al. Low temperature sweetening of higher plants. In: FRIEND, J., RODES, M.J.C. Recent advances in the biochemistry of fruits and vegetables. New York : Academic, 1981. p.41-61.
BEUKEMA, H.P., VAN DER ZAAG, D.E. Introduction to potato production. Wageningen : Centre for Agricultural Publishing Documentation (PUDOC), 1990. 208p.
CLAUSEN, A.M. Banco de Papas. Campo y Tecnologia, Balcarce, p.14-15, 1997.
COSTA, D.M. Duas novas cultivares de batata (Solanum tuberosum L.) criadas pela EMBRAPA/CNPF. Horti Sul, Pelotas, v.2, n.3, p.28-31, 1992.
CRAIG, A.L., MORRISON, I., BAIRD, E., et al. Expression of reducing sugar accumulation in interespecific somatic hybrids of potato. Plant Cell Report, v.13, p.401-405, 1994.
DAVIES, H.V., MACKAY, G.R. Exploitation of genetic variation to improve potato quality. Aspects of Applied Biology, v.39, p.45-49, 1994.
DALE, M.F.B., MACKAY, G.R. Inheritance of table and processing quality. In: BRADSAHW, J.E., MACKAY, G.R. Potato genetics. Cambridge : CAB International, 1994. p.285-315.
HAWKES, J.G., HJERTING, J.P. The potatoes of Ar gentina, Brazil, Paraguay and Uruguay. Oxford : Clarendon, 1969. 525p.
LEMOS, J. Perspectivas para a industrialização da batata. REUNIÃO TÉCNICA ANUAL DE PESQUISA E EXTENSÃO DA CULTURA DE BATATA NO RIO GRANDE DO SUL E SANTA CATARINA, 3, 1996, Santa Maria, RS. Anais... BISOGNIN, D.A. (Coord.). Santa Maria : CCR/UFSM, 1996. p.26-30.
NELSON, N.A. A photometric adaptation of Somogyi-method for determination of glucos. Journal of Biological Chemistry, Baltimore, v.153, p.375-380, 1944.
PEREIRA, A.S. Composição química, valor nutricional e industrialização. In: REIFSCHNEIDER, F.J.B. (coord.). Produção de batata. Brasília : Linha, 1987. p.12-28.
PEREIRA, A. da S., TAI, G.C.C., YADA, R.Y., et al. Potential for improvement by selection for reducing sugar content after cold storage for three potato populations. Theoretical and Applied Genetics, Viena, v.88, p.678-684, 1994.
PEREIRA, A. da S., CAMPOS, A.D. Teor de açúcares redutores em genótipos de batata (Solanum tuberosum L.). Ciência Rural, Santa Maria, v.29, n.1, p.13-16, 1999.
POPP, P.R. A industrialização de batatas no Brasil. In: BARBOSA, J.T. ENCONTRO NACIONAL DE PRODUÇÃO E ABASTECIMENTO DE BATATA, 7, 1993, Araucária. Anais... Araucária : SOB-SMAG/SMCT-IAPAR-EMATER/PR-CEASA/PR-EMBRAPA/SPSB, 1993. p.60-61.
ROCHA, B.E.G. Variabilidade isoenzimática e teor de materia seca dos clones de batata silvestre (Solanum spp.). Pelotas, 1997. 78p. Dissertação (Mestrado em Fitomelhoramento) - Curso de Pós-graduação em Agronomia, UFPEL, 1997.
SILVA, A.C.F. da. Batata: alguns aspectos importantes. Agropecuária Catarinense, Florianópolis, v.4, n.4, p.38-41, 1991.
THILL, C.A., PELOQUIN, S.J. Inheritance of potato chip color at 24-chromosome level. American Potato Journal, Orono, v.71, p.629-646, 1994.
VENCOVSKY, R., BARRIGA, P. Genética biométrica no fitomelhoramento. Ribeirão Preto : Revista Brasileira de Genética, 1992. 486 p.
Artigo original: Cienc. Rural, Fev 2001, vol.31, no.1, p.43-47. ISSN 0103-8478.
Carla Simone de Almeida Chalá1 Arione da Silva Pereira2 Angela Diniz Campos3 Judith Viégas4 Adriana Tourinho Salamoni5
angela[arroba]cpact.embrapa.br
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




