 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
O experimento foi realizado no Laboratório de Animais do Departamento de Zootecnia da Universidade Federal de Viçosa, no período de agosto a outubro de 1997. Foram utilizados quatro animais zebu, não-castrados, com peso médio inicial de 340 kg, fistulados no rúmen e no abomaso, distribuídos em um delineamento em quadrado latino 4 x 4. Os animais foram mantidos em baias individuais cobertas, de 2 x 2,5 m, com comedouros e bebedouros circulares de alvenaria, sendo pesados no início e ao final de cada período experimental.
As rações contendo feno de capim-tifton 85 de diferentes idades de rebrota foram balanceadas segundo o NRC (1996), com proporção de volumoso:concentrado de 60:40, de modo que a energia metabolizável das rações atendesse aos requerimentos dos animais para o ganho diário de 1,0 kg.
Na Tabela 1 é apresentada a composição bromatológica dos fenos e do concentrado. O feno de capim-tifton 85 foi produzido com 28, 35, 42 e 56 dias de rebrota, após uma adubação com 75 kg/ha de N, na forma de sulfato de amônio, e 60 kg/ha de K2O, como cloreto de potássio, no município de Tupaciguara, na região do Triângulo Mineiro. Na Tabela 2 encontra-se a composição percentual do concentrado usado nas quatro rações.
Consta da Tabela 3 a composição das rações experimentais. A ração foi fornecida duas vezes ao dia, às 7h30 e às 15h, exceto quando se coletaram amostras para determinação do pH e da concentração de amônia, quando se forneceu toda a ração pela manhã. A quantidade de ração fornecida diariamente foi calculada permitindo aproximadamente 10% de sobras.
Cada período experimental teve a duração de 18 dias, sendo dez dias para adaptação dos animais às rações e oito dias para coletas, dos quais quatro dias foram destinados a coletas de digesta de abomaso e fezes, para determinação das digestibilidades totais e parciais; um dia, à coleta de urina, para determinação da perda de N urinário; dois dias, à coleta de digesta ruminal, para determinação da taxa de passagem, sendo coletados também, durante o 7º dia de coletas, aproximadamente 200 mL de fluido ruminal, para determinação do pH e da concentração de N-NH3. No oitavo dia, foram coletadas amostras de fluido ruminal para o isolamento de bactérias.
As coletas de digesta do abomaso e fezes foram realizadas duas vezes ao dia, durante quatro dias, às 7h30 e 13h30, no 1º dia, às 9h e 15h, no 2º dia, às 10h30 e 16h30, no 3º dia, e às 12h e 18 h, no 4º dia (ZINN e OWENS, 1993). As porções fecais, coletadas no mesmo dia, foram armazenadas juntas, de modo que, ao final de cada período experimental, foram obtidas quatro amostras de fezes e oito de digestas de abomaso, por animal, as quais foram armazenados em freezer a aproximadamente -15ºC.
O consumo das rações foi mensurado por meio da pesagem do ofertado e das sobras, coletando-se amostras diárias da ração ofertada e das sobras, por tratamento, por animal, em cada período de coletas. Para determinação da excreção de MS fecal, utilizou-se o indicador óxido crômico, ministrado, via fístula ruminal, em duas porções diárias de 10 g cada, durante os sete dias finais do período de adaptação e os quatro dias do período de coletas, exceto à tarde do último dia.
Na ocasião do processamento, as amostras foram retiradas do freezer ao final da tarde e, no dia seguinte, levadas para uma estufa com ventilação forçada a 60-65ºC, por 72 horas. As amostras de digestas de abomaso foram pesadas, antes e após a retirada da estufa, para determinação da porcentagem de amostra seca ao ar (ASA). Após a secagem, todas as amostras foram moídas, em moinho tipo Wiley, utilizando-se peneira de 1 mm de diâmetro de poros. Aproximadamente 10 g de cada amostra seca de digesta de abomaso foram reunidos para formar uma amostra composta por animal, em cada período, sendo acondicionadas em recipientes de vidro para posteriores análises laboratoriais.
Os teores de MS, MO, PB, EE e FDN, nas amostras dos alimentos fornecidos (feno e concentrado), sobras, digestas de abomaso e fezes, além de Cr, nas duas últimas amostras, foram determinados segundo metodologias descritas por SILVA (1990).
Os carboidratos totais (CHOS) foram obtidos por meio da equação, citada por SNIFFEN et al. (1992): CHOS (%MS) = 100 - (%PB + %EE + %CINZAS). Para o cálculo do consumo de nutrientes digestíveis totais (NDTc), utilizou-se a equação, citada por SNIFFEN et al. (1992): NDTc = (PBc - PBf) + 2,25 (EEc - EEf) + (CHOSc - CHOSf), que representa os consumos e excreções fecais de proteína bruta (PBc e PBf), extrato etéreo (EEc e EEf) e carboidratos totais (CHOSc e CHOSf), respectivamente. A concentração de nutrientes digestíveis totais (NDT) foi calculada pela equação, citada por SNIFFEN et al. (1992): %NDT = (NDTc/MSc) x 100, em que MSc representa o consumo de matéria seca. A concentração de energia metabolizável (EM), em Mcal/kg, foi obtida pela equação: EM = %NDT x 4,409 x 0,82 (SNIFFEN et al., 1992).
Os dados de consumo e digestibilidades aparentes totais, ruminais e intestinais foram submetidos às análises de variância e regressão, em função da idade do feno contido na ração (28, 35, 42 e 56 dias), utilizando-se o programa SAEG versão 7.0. Os modelos foram selecionados utilizando-se como critérios a significância do coeficiente de regressão, pelo teste "t", a 1, 5 e 10% de significância, o coeficiente de determinação e o conhecimento do comportamento do fenômeno estudado.
Os resultados referentes aos consumos médios dos nutrientes, e suas respectivas equações de regressão, encontram-se na Tabela 4.
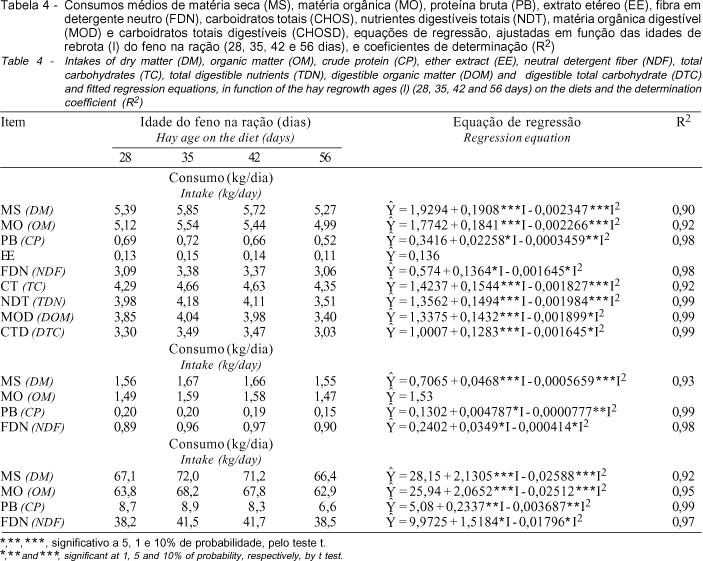
Observaram-se comportamentos quadráticos dos consumos de matéria seca e matéria orgânica (kg/dia, %PV e g/kg0,75), em função da idade do feno incluído nas dietas, exceto para o consumo de MO (%PV), que apresentou valor médio de 1,53%PV. Estimaram-se consumos máximos de MS de 5,81 kg/dia, 1,67%PV e 72 g/kg0,75, quando as rações continham feno de capim-tifton 85 de 40,6; 41,3; e 41,2 dias de rebrota, respectivamente. Estimaram-se máximos consumos de MO de 5,51 kg/dia e 68,39 g/kg0,75, quando as rações continham feno de 40,6 e 41,1 dias, respectivamente.
Em experimento com ovinos, utilizando-se, como fonte única, os mesmos fenos usados no presente experimento, ATAÍDE JR. et al. (2000) também encontraram comportamento quadrático para a ingestão de feno em função da idade de rebrota, estimando máximo consumo para feno com 39 dias de rebrota. Por outro lado, em ensaio de desempenho com bovinos da raça nelore, utilizando as mesmas rações do presente estudo, exceto aquela contendo feno de 28 dias, ATAÍDE JR. et al. (2001) verificaram comportamento linear decrescente para o consumo de MS, com reduções de 0,037 kg/dia, com o avanço da idade do feno incluído na ração. Mandebvu et al., 1998, citados por HILL et al. (1998), verificaram mais altos consumos em novilhos recebendo, como fonte única, fenos de capim-tifton 85 ou capim-coastal de plantas colhidas com 49 dias de idade do que com 21 ou 35 dias.
Os consumos de matéria seca e matéria orgânica foram aquém dos esperados, provavelmente devido ao estresse provocado pela metodologia de coletas realizada nesse tipo de experimento, uma vez que o consumo de animais nelore, em confinamento, recebendo rações semelhantes, incluindo fenos de 35 a 56 dias de rebrota, variou de 8,13 a 7,26 kg/dia (ATAÍDE JR. et al., 2001).
Os consumos de PB, expressos em kg/dia, %PV e g/kg0,75, apresentaram efeito quadrático da idade do feno na ração, estimando-se consumos máximos de PB de 0,71 kg/dia, 0,20%PV e 8,8 g/kg0,75, com a inclusão de fenos de 32,6; 30,8; e 31,7 dias de idade na ração, respectivamente, o que está consistente com os elevados teores protéicos das rações contendo fenos mais jovens. ATAÍDE JR. et al. (2001) verificaram reduções de 0,014 kg/dia, no consumo de PB, em rações contendo feno de capim-tifton 85 de 35 a 56 dias de idade. O consumo de EE, no presente trabalho, não foi influenciado pela idade do feno na ração, sendo 136 g/dia, em média.
Os consumos de FDN também foram influenciados pela idade do feno na ração, apresentando resposta quadrática, estimando-se consumos máximos de 3,4 kg/dia, 0,98%PV e 42,1 g/kg0,75, em dietas contendo feno de 41,5; 42,1; e 42,3 dias, o que se explica pelo maior consumo de MS a essas idades, visto que o teor de FDN foi similar entre as rações.
Os consumos de CHOS, NDT, MOD e CTD também apresentaram comportamentos quadráticos, encontrando-se consumos máximos de 4,69; 4,17; 4,04; e 3,50 kg/dia, respectivamente, em rações contendo fenos de 42,2; 37,6; 37,7; e 39 dias, respectivamente. Segundo MERTENS (1994), o desempenho do animal é função direta do consumo de matéria seca digerível. No presente experimento, rações contendo feno de capim-tifton 85, de 37,7 dias de rebrota, proporcionaram o mais alto consumo de MO digerível.
Os coeficientes de digestibilidade aparente total encontram-se na Tabela 5. Não se encontrou efeito da idade do feno incluído na ração sobre os coeficientes de digestibilidade aparente total de MS, MO, PB e EE, que apresentaram valores médios de 70,9; 72,6; 60,0; e 74,4%, respectivamente. Os coeficientes de digestibilidade aparente total da FDN e dos CHOS foram influenciados pela idade do feno incluído na ração, estimando-se reduções de 0,33 e 0,21 unidades percentuais por dia, respectivamente, em rações contendo feno com 28 a 56 dias de rebrota. Assim, as digestibilidades da FDN e dos CHOS variaram de 68,5 a 59,1% e de 76,9 a 71,1%, respectivamente.
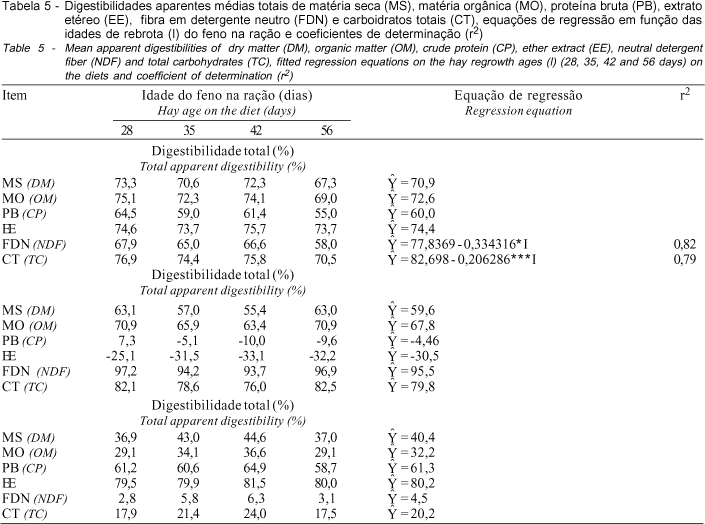
DIAS (1999), trabalhando com rações variando de 25 a 75% de concentrados, encontrou que as digestibilidades aparentes totais da MS, MO, PB, EE, FDN e CHOS, para a ração contendo 62,5% de feno de capim-coastcross, foram 53,8; 56,3, 50,8; 63,1; 56,9; e 32,4%, respectivamente. Assim, embora a formulação dos concentrados seja diferente, verifica-se que os coeficientes de digestibilidade aparente total, para rações contendo 60% de feno de capim-tifton 85, encontrados no presente trabalho, foram mais elevados do que nas rações contendo feno de capim-coastcross.
A redução na digestibilidade dos nutrientes com o avanço da idade da planta é relatada, em diferentes espécies de plantas forrageiras, por VAN SOEST (1994). Para o capim-tifton 85 e capim-coastal fornecidos como fonte única, na forma de feno, Mandebvu et al., 1998, citados por HILL et al. (1998), verificaram reduções nas digestões da FDN e da FDA, de 64,1 para 58,6% e de 58,7 para 53,1%, respectivamente, em plantas colhidas com 21 e 49 dias de idade, embora não tenham encontrado influência da idade de rebrota dos fenos sobre as digestões da MS e MO.
Em experimento com carneiros, ATAÍDE JR. et al. (2000), utilizando fenos de capim-tifton 85 semelhantes aos do presente experimento, verificaram decréscimos lineares para as digestibilidades dos nutrientes, com reduções de aproximadamente 0,16; 0,38; e 0,17 unidades percentuais, para os coeficientes de digestibilidade da MS, PB e FDN, com o aumento da idade de rebrota do feno de 28 a 56 dias. Já em experimentos com bovinos, utilizando-se as mesmas dietas do presente experimento, exceto o feno de 28 dias de rebrota, não se verificou influência da idade do feno sobre os coeficientes de digestibilidades de MS, PB, EE, FDN e CHOS (ATAÍDE JR. et al., 2001).
De modo geral, no presente trabalho, a ração contendo feno de 56 dias de idade apresentou tendência de mais baixos coeficientes de digestibilidade dos nutrientes, o que pode ser atribuído ao mais baixo teor protéico e de nutrientes digestíveis totais, além do mais alto teor de FDN indigerível do feno incluído nessa ração, como observado por HENRIQUES et al. (1998). Entretanto, verifica-se que os altos coeficientes de digestibilidade dos nutrientes encontrados para as rações contendo feno de capim-tifton 85 corrobora com vários trabalhos incluindo essa gramínea, em rações para vacas de leite e para novilhos de corte (HILL et al, 1996, 1998; WEST et al., 1995, 1998).
Não se encontrou efeito da idade do feno incluído na ração sobre a digestibilidade ruminal aparente dos nutrientes, observando-se valores médios de 59,6; 67,8; -4,36; -30,5; 95,5; e 79,8% para os coeficientes de digestibilidade ruminal aparente de MS, MO, PB, EE, FDN e CHOS, respectivamente (Tabela 5).
Observa-se que os coeficientes de digestibilidade ruminal da PB apresentaram valores negativos com o aumento da idade do feno incluído na ração, sendo que somente a dieta contendo feno de 28 dias de idade proporcionou valor médio positivo para a digestibilidade ruminal aparente da PB, de 7,3%, o que indica que houve pequena perda de N sob a forma de amônia no rúmen, pois o esperado seria um coeficiente de digestibilidade próximo de zero. Os valores médios negativos, encontrados para rações contendo fenos de 35, 42 e 56 dias de idade, indicam pequeno déficit de proteína degradada no rúmen (PDR) na ração, pois chegou ao abomaso maior quantidade de proteína do que foi ingerida. Ressalta-se que o teor protéico das dietas variou de 13,2 a 10,1%, para dietas com inclusão de fenos de 28 a 56 dias de rebrota.
Os coeficientes de digestibilidade ruminal negativos para o extrato etéreo devem-se à ocorrência de síntese de lipídeos microbianos no rúmen, o que faz com que cheguem mais lipídeos no abomaso do que a quantidade ingerida do mesmo, conforme verificado por LADEIRA (1998) e DIAS (1999).
Trabalhando com a inclusão de concentrados na ração, DIAS (1999) encontrou, para rações contendo 62,5% de feno de capim-coastcross, coeficientes de digestibilidade aparente ruminal de 57,5; 73,8; -24,8; -30,4; 138,3; e 56,9%, respectivamente, para MS, MO, PB, EE, FDN e CHOS.
Os coeficientes de digestibilidade intestinal aparente da MS, MO, PB, EE, FDN e CHOS, no presente estudo, também não foram influenciados pela idade do feno na ração, apresentando valores médios de 40,4; 32,2; 61,3; 80,2; 4,5; e 20,2%, respectivamente (Tabela 5).
Da proporção digerida no trato digestivo total, a maioria da digestão da FDN (95,5%, em média) e aproximadamente 80% dos carboidratos totais ocorreram no rúmen.
A inclusão de feno de capim-tifton 85 com 42 dias de idade, nas rações, promoveu consumo máximo dos nutrientes, excetuando-se de extrato etéreo.
As digestibilidades aparentes totais de MS, MO, PB e EE não foram influenciadas pela idade do feno na ração, enquanto as digestibilidades aparentes totais da FDN e dos CHOS decresceram 0,33 e 0,21 unidades percentuais, por dia de aumento na idade do feno na ração, respectivamente.
As digestibilidades aparentes ruminais e intestinais não foram influenciadas pela idade do feno na ração.
Agradecimento
Aos irmãos Marcos e César Prado, proprietários da Fazendas Prado, em Tupaciguara, MG, pelo fornecimento dos fenos utilizados neste trabalho.
ATAÍDE JR., J.R., PEREIRA, O.G., GARCIA, R. et al. 2000. Valor nutritivo do feno de capim-tifton 85 (Cynodon spp.) em diferentes idades de rebrota, em ovinos. Rev. bras. zootec., 29(6):2193-2199 (suplemento 2).
ATAÍDE JR., J.R., PEREIRA, O.G., VALADARES FILHO, S.C. et al. 2001. Consumo, digestibilidade e desempenho de novilhos alimentados com rações à base de feno de capim-tifton 85, em diferentes idades de rebrota. Rev. bras. zootec., 30(1):215-221.
BURTON, G.W., GATES, R.N., HILL, G.M. 1993. Registration of "tifton 85" bermudagrass. Crop Sci., 33(3):644-645.
CARVALHO, A.U., VALADARES FILHO, S.C., COELHO DA SILVA, J.F. et al. Efeito dos níveis de concentrados sobre a digestão parcial dos nutrientes em zebuínos. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 33, Fortaleza, 1996. Anais... Fortaleza: SBZ, 1996. p.64-66.
CONRAD, H.R. 1966. Symposium on factors influencing the voluntary intake of herbage by ruminants: physiological and physical factors limiting feed intake. J. Anim. Sci., 25: 227-235. [
Medline ]DIAS, H.L.C. Consumo, digestibilidade e eficiências microbiana em novilhos F1 limousin x nelore alimentados com dietas contendo cinco níveis de concentrado. Viçosa, MG, UFV, 1999. 76p. Tese (Mestrado em Zootecnia), Universidade Federal de Viçosa, 1999.
HENRIQUES, L.T., PEREIRA, O.G., VALADARES FILHO, S.C. et al. Degradabilidade "in situ" da matéria seca e da fibra em detergente neutro do feno de tifton-85 (Cynodon spp.), em quatro idades de rebrota. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 35, Botucatu, 1998. Anais....Botucatu, SBZ, 1998. p.570-572.
HATFIELD, R.D., MANDEBVU, P., WEST, J. 1997. A comparison of tifton 85 and coastal bermudagrass cell walls. (
http://www.dfrc.ars.usda.gov./Research_Sumaries/1997).HILL, G.M., GATES, R.N., BURTON, G.W. 1993. Forage quality and grazing steer performance from tifton 85 and tifton 78 bermudagrass pastures. J. Anim. Sci., 71(12):3219-3225. [
Medline ]HILL, G.M., GATES, R.N., WEST, J.W. et al. Tifton 85 bermudagrass utilization in beef, dairy, and hay production. In: WORKSHOP SOBRE O POTENCIAL FORRAGEIRO DO GÊNERO CYNODON, 1996, Juiz de Fora. Anais... Juiz de Fora: EMBRAPA - CNPGL, 1996. p. 139-150.
HILL, G.M., GATES, R.N., WEST, J.W. et al. Pesquisa com capim bermuda cv. Tifton 85 em ensaios de pastejo e de digestibilidade de feno com bovinos. In: PEIXOTO, A.M.; MOURA, J.C.; FARIA, V.P. SIMPÓSIO DO MANEJO DE PASTAGENS DE TIFTON, COAST-CROSS E ESTRELA, 1998, Piracicaba. Anais... Piracicaba: ESALQ, 1998. p. 7-22.
HILL, G.M., GATES, R.N., WEST, J.W. et al. 1997a. Bermudagrass cultivar maturity effects on hay digestibility in steers. J. Anim. Sci., 75:201.
HILL, G.M., GATES, R.N., WEST, J.W. et al. 1997b. Consumption and digestibility of Coastal, tifton 78, and tifton 85 hays harvested at two maturity stages when fed to steers. (
http://www.ads.uga.edu/annrpt/1997/97_031.htm).LADEIRA, M.M., VALADARES FILHO, S.C., LEÃO, M.I. et al. Dietas contendo diferentes níveis de concentrado em novilhos nelore: digestibilidade aparente parcial. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 35, Botucatu, 1998. Anais... Botucatu, SBZ, 1998. p.579-581.
MERTENS, D.R. Análise da fibra e sua utilização na avaliação e formulação de rações. In: SIMPÓSIO INTERNACIONAL DE RUMINANTES, REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 29, Lavras, 1992. Anais... Lavras, SBZ, 1992. p.188-219.
MERTENS, D.R. 1994. Regulation of forage intake. In: FAHEY JR., G.C., COLLINS, M., MERTENS, D.R., MOSER, L.E. (Eds). Forage quality, evaluation and utilization. Madison, Wisconsin: ASA-CSSA-SSSA. p.450-493.
MERTENS, D.R., ELY, L.O. 1979. A dynamic model of fiber digestion and passage in the ruminant for evaluating forage quality. J. Anim. Sci., 49(4):1085-1095.
NATIONAL RESEARCH COUNCIL - NRC. 1996. Nutrient requirements of beef cattle. 7.ed. Washington, D.C.: National Academy. 242p.
SILVA, D.J. 1990. Análise de alimentos (Métodos químicos e biológicos). Viçosa: UFV. 165p.
SNIFFEN, C.J., O'CONNOR, J.D., VAN SOEST, P.J. et al. 1992. A net carbohydrate and protein system for evaluating cattle diets: II. Carbohydrate and protein availability. J. Anim. Sci., 70(11):3562-3577. [
Medline ]VAN SOEST, P.J. 1965. Symposium on factors influencing the voluntary intake of herbage by ruminants: Voluntary intake relation to chemical composition and digestibility. J. Anim. Sci., 24(3):834-844
VAN SOEST, P.J. 1994. Nutritional ecology of the ruminant. Ithaca, New York: Cornell. 476p.
WALDO, D.R., JORGENSEN, N.S. 1981. Forages for high animal production: nutritional factors and effects of conservation. J. Dairy Sci., 64:1207-1229.
WEST, J.W., MANDEBVU, P., HILL, G.M. et al. 1998. Intake, milk yield, and digestion by dairy cows fed diets with incresing fiber content from bermudagrass hay or silage. J. Dairy Sci., 81:1599-1607. [
Medline ]WEST, J.W., MATHIS, M.J., HILL, G.M. et al. 1995. Effects of dietary forage source and fiber content on intake and lactational performance of dairy cows. (
http://www.ads.uga.edu/annrpt/1995/95_186.htm).ZINN, R.A., OWENS, F.N. 1993. Ruminal escape protein for lightweight feedlot calves. J. Anim. Sci., 71:1677-1687. [
Medline ]Karina Guimarães Ribeiro2, Rasmo Garcia3, Odilon Gomes Pereira3, Sebastião de Campos Valadares Filho3, Paulo Roberto Cecon4
rgarcia[arroba]ufv.br Parte da tese de Doutorado do primeiro autor, parcialmente financiada pela FAPEMIG (CAG 2316/96).2
Professora da FESURV, Rio Verde - GO. Campus Universitário, Cx. Postal 104, Cep 75901-970, Rio Verde, GO.3
Professores do DZO/UFV, Viçosa - MG, Bolsitas do CNPq.4
Professor do DPI/UFV, Viçosa - MG, Bolsista do CNPq.| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|







