 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
Os insetos foram coletados em campo e mantidos em recipientes plásticos, por pelo menos sete dias, alimentados com ração esterilizada para aves e em condições ambiente (24 ± 2°C). Foram avaliados 99 isolados de B. bassiana e M. anisopliae, provenientes das coleções do Instituto Biológico de São Paulo, da Universidade Estadual de Londrina e da Unioeste, mantidos na forma de conídios em freezer (-10°C).
Avaliação da patogenicidade. As culturas fúngicas foram obtidas por multiplicação em placas de Petri contendo meio de cultura BDA ou meio para produção de esporos (Alves et al. 1998), incubados em câmara BOD (26 ± 1°C, 14h de fotofase), por um período de sete a dez dias para crescimento vegetativo e conidiogênese. Após este período, os conídios foram coletados, raspando-se o meio de cultura e armazenados em tubos de vidro para posterior utilização.
Nos bioensaios, larvas com 1 cm de comprimento e adultos foram transferidos para recipientes plásticos descartáveis onde foram imersos em 1 ml de suspensão de conídios na concentração de 109 conídios viáveis/ml (água + espalhante adesivo Tween® 80 - 0,01%), sendo agitados manualmente durante 10 segundos. Na testemunha, os insetos foram imersos em solução aquosa de espalhante adesivo Tween® 80 (0,01%).
Em seguida, as larvas foram individualizadas em caixas de acrílico (anulando o comportamento canibal) e os adultos mantidos em placas de Petri, ambas forradas com papel-filtro e contendo ração estéril para aves. Os recipientes foram mantidos em condições controladas, em recipientes plásticos contendo espuma umedecida em uma câmara BOD (26 ± 1°C e 14h de fotofase), e avaliados diariamente, durante dez dias. Neste período, a água foi reposta na espuma para manutenção da umidade. Para cada isolado, foram utilizadas quatro repetições com 15 insetos. Diariamente, durante 10 dias, foi avaliada a mortalidade sendo os cadáveres imersos em solução de álcool 70% e água destilada, e transferidos individualmente para câmara úmida para permitir o desenvolvimento do fungo, visando à confirmação do agente causal da doença.
Estimativa da virulência. Estimou-se a CL50 dos isolados que provocaram mortalidade confirmada acima de 80% para larvas e 50% para adultos, sendo avaliado também crescimento vegetativo, produção de conídios em meio de esporulação (ME e arroz e em cadáveres de larvas e adultos.
Foram preparadas diluições seriadas, padronizadas nas concentrações de 105, 106, 107, 108 e 109 conídios viáveis/ml. Os bioensaios foram realizados utilizando-se o mesmo procedimento, sendo que para efeito de cálculos, considerou-se a mortalidade confirmada observada no 5º e no 10º dia após inoculação do fungo.
Os dados foram submetidos à análise de Probit (SAS Institute Inc. 1995), porém, devido ao não ajuste dos dados ao modelo estabelecido, procedeu-se à análise de variância (teste F) e as médias foram comparadas entre si pelo teste de Tukey (P < 0,05), segundo o delineamento experimental inteiramente casualizado e delineamento fatorial.
Procurou-se determinar eventuais interações entre a concentração de conídios nas suspensões e os diferentes isolados quanto à mortalidade de larvas e adultos de cascudinho, utilizando-se o programa estatístico Sisvar (Ferreira 2000), usando dados transformados em arcsen ![]() , com o auxílio do programa Minitab®.
, com o auxílio do programa Minitab®.
Crescimento vegetativo. Os isolados foram inoculados em três pontos na superfície do ME em placas de Petri, e incubados a 26 ± 1°C e 14h de fotofase. Após 10 dias, avaliou-se o tamanho das colônias fazendo-se duas medições perpendiculares, determinando-se o diâmetro médio. Para cada isolado foram preparadas três placas, sendo cada colônia considerada uma repetição. Os dados foram submetidos à análise de variância (teste F) e as médias comparadas entre si pelo teste de Tukey (P < 0,05), segundo o delineamento experimental inteiramente casualizado, sendo que os dados foram transformados em ![]() utilizando-se o programa estatístico Sisvar (Ferreira 2000).
utilizando-se o programa estatístico Sisvar (Ferreira 2000).
Produção de conídios em meio de cultura. Após a avaliação do crescimento vegetativo, as colônias foram recortadas do meio de cultura, na linha terminal do halo da colônia e individualizadas em tubos de vidro de fundo chato com 10 ml de água destilada e Tween 80 a 0,01%. Os conídios foram removidos com o auxílio de um pincel, submetendo-se, em seguida, os tubos a ação de vórtex por um minuto. A quantificação dos conídios foi feita em câmara de Neubauer e a análise dos dados foi executada com base nos procedimentos descritos para o experimento de avaliação do crescimento vegetativo.
Produção de conídios em arroz. A avaliação da produção de conídios em arroz foi realizada com os isolados UNIOESTE 02 e UNIOESTE 04, utilizando o método do Biomax® (Alves & Pereira 1989). Para tal, sacos de polipropileno contendo 200 g de arroz polido e pré-cozido por três minutos em água fervente, foram autoclavados por 15 min a 120°C. Após o resfriamento, foi realizada a inoculação (10 ml de suspensão de conídios produzidos em meio de esporulação), sendo que para cada isolado foram preparadas três repetições, com 10 sacos para cada uma.
Os sacos foram incubados a 26 ± 1°C e 14h de fotofase durante sete dias, quando foram abertos e seu conteúdo exposto a um fluxo de ar em bandejas de plástico, em uma câmara asséptica por quatro dias, sob temperatura de 25 ± 1°C e 24h de fotofase. Em seguida efetuou-se a avaliação da produção de conídios/g e a viabilidade, seguindo procedimentos descritos por Leite et al. (2003).
Produção de conídios em cadáveres de cascudinho. Os isolados previamente selecionados e multiplicados foram inoculados em larvas e adultos de cascudinho, conforme metodologia descrita no teste de patogenicidade. Foram selecionados cinco larvas e cinco adultos mortos pelo fungo, tendo como critério de seleção a plena conidiogênese, sendo cada um dos lotes de cadáveres considerados uma repetição (total de 3 repetições).
Em seguida, os conídios da superfície dos insetos foram removidos imergindo-os separadamente cada uma das repetições em 10 ml de água destilada com espalhante adesivo Tween 80 (0,01%). A suspensão foi agitada em vórtex durante um minuto e quantificada (câmara de Neubauer). Os dados foram submetidos à análise de variância (teste F) e as médias comparadas entre si pelo teste de Tukey, ambos a 5% de significância, segundo o delineamento experimental inteiramente casualizado e o delineamento fatorial, verificando-se eventuais interações entre os estágios de desenvolvimento e os diferentes isolados quanto à produção de conídios em cadáveres, sendo que os dados foram transformados em utilizando-se o programa estatístico Sisvar (Ferreira 1992).
Avaliação da patogenicidade. Os isolados de B. bassiana foram mais eficientes contra o cascudinho em relação aos isolados de M. anisopliae, sendo que a mortalidade larval confirmada variou entre 6,7 e 100% e a mortalidade confirmada de adultos variou entre 0 e 86,7%. Dentre os 58 isolados de B. bassiana, 18 causaram mortalidade larval confirmada superior a 80% e sete entre 50% e 80% para os adultos (Tabela 1).

Para M. anisopliae, a mortalidade larval confirmada variou entre 8,3% e 90% e mortalidade confirmada de adultos variou entre 0 e 90%. No entanto, somente seis isolados de um total de 41 causaram mortalidade larval confirmada superior a 80% e apenas dois deles causaram mortalidade de adultos entre 50% e 80% (Tabela 2).
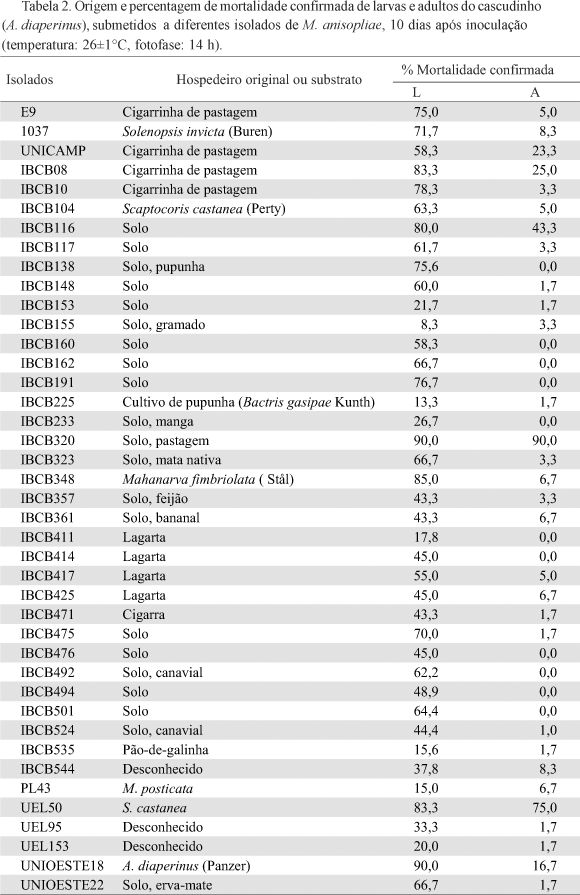
Assim, foram selecionados para os estudos posteriores, cincos isolados de B. bassiana, (UEL 25, UNIOESTE 02, UNIOESTE 04, UNIOESTE 05 e UNIOESTE 40) e um isolado de M. anisopliae (IBCB 320).
Embora o isolado UNIOESTE 40 tenha causado mortalidade larval de 55%, ele foi selecionado por ter atingido um índice elevado de mortalidade de adultos (78,3%). Por outro lado, o isolado UEL 50 (M. anisopliae) não foi selecionado apesar de inicialmente ter apresentado 83,3% de mortalidade larval e 75% de mortalidade de adultos, pois esses níveis de mortalidade não foram confirmados nos estudos posteriores.
Dentre os isolados mais efetivos, encontram-se aqueles obtidos de cadáveres de cascudinhos (UNIOESTE 02, UNIOESTE 04 e UNIOESTE 05), sendo que resultados semelhantes foram obtidos por Geden et al. (1998), que testando vários isolados de B. bassiana, também verificaram que os mais virulentos foram isolados do cascudinho.
Neste estudo, observou-se ainda que o estágio larval foi mais suscetível quando comparado ao estágio adulto, sendo que 11 isolados de M. anisopliae e sete de B. bassiana não causaram mortalidade em adultos. Steinkraus et al. (1991) constataram que larvas de cascudinhos foram mais suscetíveis à infecção por B. bassiana. Da mesma forma, a maior suscetibilidade das larvas foi também observada por Geden et al. (1998) e por Crawford et al. (1998). Esta diferença pode estar relacionada a diferenças no exoesqueleto nos dois estágios de desenvolvimento. Assim, os adultos apresentam o tegumento mais esclerotizado do que as larvas dificultando a penetração do patógeno.
No Brasil, Alexandre et al. (2006), comparando a eficiência de B. bassiana e M. anisopliae em diferentes temperaturas, constataram diferenças significativas entre a suscetibilidade de larvas e adultos de cascudinho, sendo que a mortalidade larval variou entre 77% e 95% a 26ºC e entre 2% e 68% a 32ºC, e a mortalidade de adultos variou entre 0 e 82% a 26ºC e 0 e 26% a 32ºC.
Vários estudos ressaltam a eficiência de isolados de B. bassiana no controle do cascudinho, indicando ser este um fungo promissor no controle da praga (Steinkraus et al. 1991). Esses autores observaram que B. bassiana é muito freqüente nas populações de cascudinho em aviários nos EUA. A associação também foi constatada no Brasil, por Alves et al. (2005), que observaram elevado índice de controle natural sobre larvas e adultos nas áreas externas do aviário. Também a ocorrência de M. anisopliae foi relatada em populações de cascudinho em um aviário comercial em Cascavel, PR (Alves et al. 2004).
Por outro lado, em condições de laboratório, Chernaki-Leffer (2004) observou maior porcentagem de mortalidade de A. diaperinus (40%) com isolados de M. anisopliae, em relação a Beauveria sp. e Paecilomyces sp. Nesse estudo, todos os isolados apresentaram elevada CL50, indicando reduzida virulência dos mesmos. Essa variação observada, contrastante com os resultados aqui obtidos, provavelmente está relacionada à utilização de métodos diferentes de inoculação além de diferentes isolados.
Estimativa da virulência. Os dados obtidos não se ajustaram ao modelo Probit, provavelmente porque os bioensaios nem sempre seguiram o modelo do tipo estímulo-resposta (Haddad 1998).
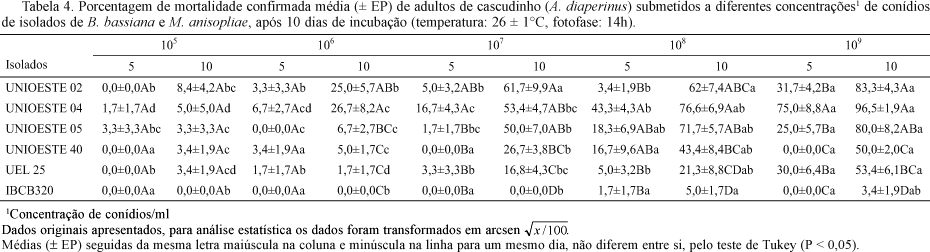
Os valores de mortalidade confirmada nas diferentes concentrações de conídios apresentaram diferenças significativas tanto entre os isolados como entre espécies de fungo, além de diferenças entre os estágios de desenvolvimento do inseto (Tabelas 3 e 4).
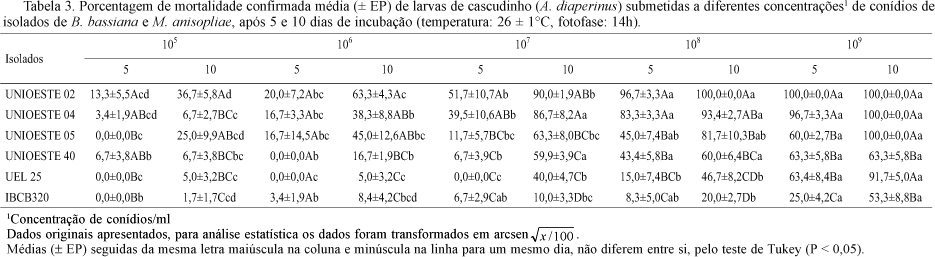
Nas concentrações 105 e 106 conídios/ml, todos os isolados causaram baixa mortalidade em larvas e adultos, após o 5º dia da inoculação. Porém, no 10º dia após inoculação, somente a mortalidade de adultos foi baixa para todos os isolados, variando entre 0 e 25%. Para larvas, na concentração 105conídios/ml, os níveis de mortalidade também foram baixos, no entanto, ocorreram diferenças significativas entre os isolados, variando de 1,7% a 36,7%, sendo os maiores valores obtidos para UNIOESTE 02 e os menores para IBCB 320. Na concentração 106 a mortalidade larval variou entre 5% e 63%, novamente tendo destaque o isolado UNIOESTE 02.
Em todas as concentrações, após o 5º dia, os isolados UNIOESTE 02 e UNIOESTE 04 apresentaram os maiores níveis de mortalidade destacando-se dos demais. A superioridade do isolado UNIOESTE 02 manteve-se para larvas, enquanto que para adultos, o melhor isolado foi UNIOESTE 04.
Nas concentrações 107, 108 e 109conídios/ml, no 10º após inoculação, os isolados de B. bassiana provenientes de cadáveres do inseto (UNIOESTE 02, UNIOESTE 04 e UNIOESTE 05) causaram maior mortalidade confirmada, tanto em larvas como em adultos, sendo que, na concentração mais elevada, eles causaram 100% de mortalidade confirmada de larvas.
Esses resultados concordam com os relatados por Geden et al. (1998). Os autores avaliaram em laboratório a virulência de quatro isolados de B. bassiana sobre todos os estágios de desenvolvimento do cascudinho e verificaram que aqueles obtidos de cadáveres do inseto foram mais eficientes para o seu controle em relação aos obtidos em cadáveres de Musca domestica (L.).
Os isolados UNIOESTE 40 e UEL 25 de B. bassiana, apresentaram valores de mortalidade intermediários tanto para larvas quanto para adultos, causando na maior concentração (109), 63,3% e 91,7% de mortalidade larval e 50% e 53,4% de mortalidade de adultos, respectivamente.
O isolado de M. anisopliae testado nessa fase do estudo (IBCB 320) foi menos virulento quando comparado aos demais, sendo que nas concentrações abaixo de 109, matou menos que 25% da população de larvas e, somente na maior concentração esteve entre os valores intermediários com 53,3% de mortalidade larval. Para adultos causou mortalidade apenas nas concentrações 108 e 109 conídios/ml e ainda assim, inexpressivas (5% e 3,4%, respectivamente).
Assim, a superioridade de B. bassiana em relação a M. anisopliae foi confirmada também nos experimentos de virulência, indicando ter esta espécie maior potencial de uso no controle do cascudinho em aviários.
Crescimento vegetativo e produção de conídios em meio de cultura. Em relação ao crescimento vegetativo, UNIOESTE 02 foi o que teve maior diâmetro médio da colônia com 4,3 cm e o isolado UNIOESTE 40 o menor, com média de 2,9 cm, diferença de cerca de 1,5 vezes entre o maior e o menor diâmetro (Tabela 5). Os demais isolados de B. bassiana (UNIOESTE 04, UNIOESTE 05, UEL 25) apresentaram valores de crescimento intermediários, variando de 3,6 cm a 3,9 cm.
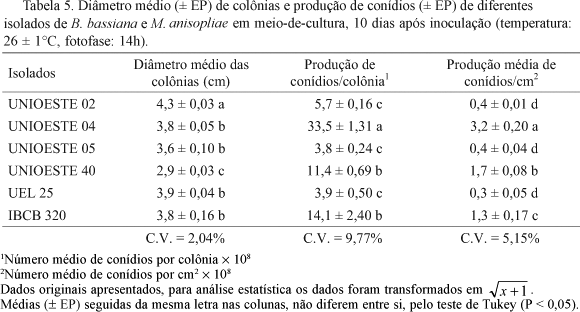
Valores próximos aos aqui obtidos foram observados por Batista Filho et al. (2001) com isolado de B. bassiana (4,3 cm) e M. anisopliae (3,5 cm). Entretanto, Oliveira & Neves (2004) verificaram que B. bassiana apresentou colônias com diâmetro médio de 2,6 cm após oito dias de incubação.
Ocorreram variações significativas em relação à produção de conídios em meio de cultura, sendo que UNIOESTE 04 teve destaque em relação aos demais isolados analisados, independente de se analisar produção de conídios/cm2 (3,2 ´ 108) ou conídios/colônia (33,5 ´ 108). Os menores valores corresponderam aos dos isolados UNIOESTE 02, UEL 25 e UNIOESTE 05, mantendo esta inferioridade tanto em produção/cm2 como em colônia (Tabela 5).
Valores próximos aos do presente trabalho foram apresentados por Batista Filho et al. (2001), que obtiveram produção de 9,8 ´ 108conídios/colônia para B. bassiana e 35,1 ´ 108conídios/colônia para M. anisopliae.
Comparando os parâmetros de crescimento vegetativo com os de produção de conídios, não foi observada qualquer relação direta entre eles, já que o isolado UNIOESTE 02, que apresentou o maior diâmetro, foi um dos menos produtivos e, os isolados UNIOESTE 04, UNIOESTE 05, UEL 25 e IBCB 320, que estiveram entre os isolados de crescimento intermediário, variaram quanto à produção de conídios, UNIOESTE 04 foi o mais produtivo, IBCB 320 de produção intermediária e os isolados UNIOESTE 05 e UEL 25 apresentaram um dos valores mais baixos de produção. O isolado UNIOESTE 40 que teve o menor crescimento vegetativo, apresentou produção intermediária de conídios (Tabela 5).
Produção de conídios em arroz. O isolado UNIOESTE 04 manteve elevada produção, apresentando uma média de 8,3 ´ 108conídios/g arroz, com 85% de conídios viáveis. Resultado inferior ao observado por Leite et al. (2003), que foi de 6 ´ 109conídios/g de arroz. Por outro lado, embora o isolado UNIOESTE 02 tenha apresentado formação do micélio, não produziu conídios.
Produção de conídios em cadáveres de cascudinho. Houve variações significativas na produção de conídios em cadáveres entre os diferentes estágios de desenvolvimento do inseto e nos diferentes isolados. O isolado UNIOESTE 04 teve a maior produção de conídios em larvas, sendo cerca de 150 vezes mais produtivo em relação ao isolado UEL 25. Em adultos, a diferença foi de aproximadamente 58 vezes entre o isolado mais produtivo (UNIOESTE 04) e o menos produtivo (UNIOESTE 05) (Tabela 6).

No entanto, alguns isolados, tais como UNIOESTE 02 e IBCB 320 não apresentaram plena conidiogênese sobre os cadáveres, tendo sido observada a presença de micélio e conídios sobre os mesmos, confirmando a mortalidade pelo fungo, embora não sendo suficiente para permitir a quantificação segura dos conídios.
A superioridade do isolado UNIOESTE 04, verificada anteriormente na produção de conídios em placa e em arroz, manteve-se também em produção de conídios em cadáveres, tanto em larvas (30,3 ´ 106 conídios/inseto) como em adultos (21 ´ 106 conídios/inseto), não havendo diferenças significativas entre os dois estágios de desenvolvimento.
O isolado IBCB 320 apresentou a segunda maior produção de conídios em larvas (26,33 ´ 106 conídios/inseto). Porém, não apresentou mortalidade confirmada em adultos, não permitindo a determinação de produção de conídios nesse estágio de desenvolvimento.
Os isolados UNIOESTE 05, UNIOESTE 40 e UEL 25 apresentaram baixas produções de conídios em larvas, não diferindo estatisticamente entre si. Por outro lado, em adultos os isolados UNIOESTE 40 e UEL 25 tiveram destaque com 9 ´ 106 e 4,8 ´ 106 conídios/inseto, respectivamente, enquanto que UNIOESTE 05 teve a menor produção em adultos com 0,4 ´ 106 conídios/inseto.
Alexandre et al. (2006), avaliando a conidiogênese de isolados de B. bassiana UNIOESTE 02 e UEL 25 sobre o cascudinho, obtiveram maior produção em relação aos resultados aqui apresentados. A diferença pode estar relacionada ao fato de que no presente trabalho utilizou-se uma suspensão de conídios com a concentração cerca de dez vezes maior que a concentração de conídios utilizada pelos referidos autores. Isto pode ter conduzido a maior competição devido à baixa disponibilidade de nutrientes.
Neste sentido, Luz et al. (1999), avaliando a conidiogênese de B. bassiana em cadáveres de Triatoma infestans (Klug.) (Hemiptera: Reduviidae) obtiveram maior produção de conídios sobre os cadáveres expostos às concentrações menores de fungo. Resultados semelhantes foram observados por Tafera & Pringle (2003), testando o efeito de B. bassiana sobre Chilo partellus (Swinhoe) (Lepidoptera: Crambidae).
No entanto, a relação observada não segue um padrão, já que Coutinho & Oliveira (1991), testando a produção de conídios de um isolado de B. bassiana sobre cadáveres de Anthonomus grandis (Boh.) (Coleoptera: Curculionidae) verificaram que, com o aumento da concentração de conídios, ocorreu um aumento na produção de conídios sobre os cadáveres, variando entre 6,59 ´ 106 e 2,26 ´ 107 conídios/inseto para as concentrações de 3,72 ´ 107 e 3,72 ´ 1010.
Assim, considerando que os isolados UNIOESTE 02 e UNIOESTE 04 foram os que apresentaram melhor desempenho, verificou-se que não houve relação direta entre os parâmetros avaliados, já que UNIOESTE 02 esteve entre os isolados que apresentaram o melhor desempenho em relação à mortalidade de larvas e adultos, porém destacou-se somente no crescimento vegetativo, e na produção de conídios em meio de cultura esteve entre os piores (0,4 ´ 108conídios/cm2), e em cadáveres, não apresentou conidiogênese suficiente para contagem. Assim, este isolado pode não ser indicado para utilização em um programa de controle do cascudinho.
Por outro lado, o isolado UNIOESTE 04, além de ter provocado elevada mortalidade confirmada tanto para larvas como para adultos (100% e 96,5%, respectivamente na maior concentração), apresentou uma das maiores médias de crescimento vegetativo (diâmetro igual a 3,8 cm), e principalmente maior produção de conídios em placa (3,2 ´ 108conídios/cm2) e em arroz (8,3 ´ 108 conídios/g de arroz). Além disso, esse isolado foi o que produziu maior número de conídios em cadáveres, tanto em larvas (30,3 ´ 106 conídios/cadáver) como em adultos (21 ´ 106 conídios/cadáver). Esse parâmetro pode ser determinante na ocorrência de epizootias com reflexo direto no controle do inseto em campo, com relação à transmissão de insetos mortos para inseto sadios. Portanto, o isolado UNIOESTE 04 apresentou o maior potencial para utilização a campo na estratégia de manejo do cascudinho. Entretanto, são ainda necessários mais estudos para definir de forma mais precisa a incorporação deste agente de controle no manejo do cascudinho.
Aos estagiários do Laboratório de Zoologia/Unioeste/CCBS, pelo auxílio nas avaliações e ao CNPq pela concessão de Bolsa de Iniciação Científica e de Produtividade em Pesquisa.
Alexandre, T.M., L.F.A. Alves, P.M.O.J. Neves & S.B. Alves. 2006. Efeito da temperatura e substrato sobre Beauveria bassiana e Metarhizium anisopliae e sua relação no controle do cascudinho (Alphitobius diaperinus) (Panzer) (Coleoptera: Tenebrionidae). Neotrop. Entomol. 35: 75-82.
Alves, L.F.A., M.H. Gassen, F.G.S. Pinto, P.M.O. J. Neves & S.B. Alves. 2005. Ocorrência natural de Beauveria bassiana (Bals.) Vuilleman (Moniliales: Moniliaceae) sobre o cascudinho (Alphitobius diaperinus) (Panzer) (Coleoptera: Tenebrionidae), em aviários comerciais de Cascavel, PR. Neotrop. Entomol. 34: 507-510.
Alves, L.F.A., V.S. Alves, D.F. Bressan, P.M.O. J. Neves & S.B. Alves. 2004. Ocorrência de Metarhizium anisopliae (Metsch.) Sorok. (Moniliales: Moniliaceae) em adultos de cascudinho (Alphitobius diaperinus) (Panzer) (Coleoptera: Tenebrionidae), em aviários comerciais em Cascavel, PR, Brasil. Neotrop. Entomol. 33: 793-795.
Alves, S.B., J.E.M. Almeida, A. Moino Jr. & L.F.A. Alves. 1998. Técnicas de laboratório, p. 21-38. In S.B. Alves, Controle microbiano de insetos, Piracicaba, Fealq, 2ed., 1163p.
Alves, S.B. & R.W. Pereira.1989. Produção de Metarhizium anisopliae (Metsch.) Sorok. e Beauveria bassiana (Bals.) Vuill. em bandejas. Ecossistema 14: 188-192.
Arends, J.J. 1987. Control, management of the litter beetle. Poult. Digest 30: 172-176.
Batista Filho, A.B., J.E.M. Almeida & C. Lamas. 2001. Effect of thiamethoxan on entomopathogenic microorganisms. Neotrop. Entomol. 30: 437-447.
Castrillo, L.A., B.M. Wiegmann & W.M. Brooks. 1999. Genetic variation in Beauveria bassiana populations associated with the darkling beetle, Alphitobius diaperinus. J. Invertebr. Pathol. 73: 269-275.
Castrillo, L.A. & W.M. Brooks. 1998. Differentiation of Beauveria bassiana isolates from the darkling beetle, Alphitobius diaperinus, using isozyme and RAPD analyses. J. Invertebr. Pathol. 72: 190-196.
Chernaki-Leffer, A.M. 2004. Dinâmica populacional, estimativa da resistência a inseticidas e suscetibilidade do cascudinho Alphitobius diaperinus (Panzer, 1797) (Coleoptera: Tenebrionidae) a inseticidas reguladores de crescimento e a fungos entomopatogênicos. Tese de doutorado, Universidade Federal do Paraná, Curitiba, 62p.
Chernaki-Leffer, A.M., S.M. Biesdorf, L.M. Almeida, E.V.B. Leffer & F. Vigne. 2002. Isolamento de enterobactérias em Alphitobius diaperinus e na cama de aviários no Oeste do estado do Paraná, Brasil. Rev. Bras. Avícola 4: 243-247.
Coutinho, J.L.B. & J.V. Oliveira. 1991. Patogenicidade do isolado I-149Bb de Beauveria bassiana (Bals.) Vuill. a adulto de Anthonomus grandis (Coleoptera: Curculionidae). An. Soc. Entomol. Brasil 1: 199-207.
Crawford, P.J., W.M. Brooks & J.J. Arends. 1998. Efficacy of field-isolated strains of Beauveria bassiana (Moniliales: Moniliaceae) as microbial control agents of the lesser mealworm (Coleoptera: Tenebrionidae). J. Econ. Entomol. 91: 1295-1301.
Despins, J.L. & R.C. Axtell. 1994. Feeding behavior and growth of turkey pouts fed larvae of the darkling beetle, Alphitobius diaperinus. Poult. Sci. 73: 1526-1533.
Despins, J.L. & R.C. Axtell. 1995. Feeding behavior and growth of broiler chicks fed larvae of the darkling beetle, Alphitobius diaperinus. Poult. Sci. 74: 331-336.
Ferreira, D.F. 1992. SISVAR (Sistema para análise de variância para dados balanceados). Lavras, UFLA, 79p.
Geden, C.J. 1989. Understanding and controlling litter beetles. Poult. Digest, march, p.142-144.
Geden, C.J., J.J. Arends, D.A. Ruttz & D.C. Steinkraus. 1998. Laboratory evaluation of Beauveria bassiana (Moniliales: Moniliaceae) against the lesser mealworm, Alphitobius diaperinus (Coleoptera: Tenebrionidae), in poultry litter, soil and a pupal trap. Biol. Control. 13: 71-77.
Goodwin, M.A. & W.D. Waltman. 1996. Transmission of Eimeria, viruses, and bacteria to chicks: Darkling beetles (Alphitobius diaperinus) as vector of pathogens. J. Appl. Poult. Res. 5: 51-55.
Haddad, M.L. 1998. Utilização do Polo-PC para análise de Probit, p.21-38. In S.B. Alves, Controle microbiano de insetos, Piracicaba, Fealq, 2ed., 1163p.
Leite, L.G., A. Batista Filho, J.E.M. Almeida & S.B. Alves. 2003. Processos de produção, p. 33-44. In L.G. Leite, A. Batista Filho, J.E.M. Almeida & S.B. Alves. Produção de fungos entomopatogênicos. Ribeirão Preto, Alexandre de Sene Pinto, 92p.
Luz, C., M.S. Tigano, L.G. Silva & C.M. Cordeiro. 1999. Sporulation of Beauveria bassiana on cadavers of Triatoma infestans after infection at different temperatures and doses of inoculum. Mem. Inst. Oswaldo Cruz. 73: 223-225.
Mcallister, J.C., C.D. Steelman, L.A. Newberry, J.K. Skeeles. 1995. Isolation of infectious bursal disease virus from the lesser mealworm, Alphitobius diaperinus (Panzer). Poult. Sci. 74: 45-49.
Oliveira, R.C. & P.M.O.J. Neves. 2004. Compatibility of Beauveria bassiana with acaricides. Neotrop. Entomol. 33: 353-358.
Pereira, R.M., S.B. Alves & P.R. Reis. 1998. Segurança no emprego de entomopatógenos, p.21-38. In S.B. Alves, Controle microbiano de insetos, Piracicaba, Fealq, 2ed., 1163p.
SAS Institute. 1995. SAS user's guide: statistics. Cary, N.C., SAS Institute Inc.
Steinkraus, D.C., C.J. Geden & D.A. Rutz. 1991. Susceptibility of lesser mealworm (Coleoptera: Tenebrionidae) to Beauveria bassiana: Effects of host stage, formulation, substrate and host passage. J. Med. Entomol. 28: 314-321.
Tafera, T. & K.L. Pringle. 2003. Effect of exposure method to Beauveria bassiana and conidia concentration on mortality, mycosis, and sporulation in cadavers of Chilo partellus (Lepidoptera: Pyralidae). J. Invertebr. Pathol. 84: 90-95.
Watson, D.W., J.S. Guy. & S.M. Strincham. 2000. Limited transmission of turkey coronavirus in young turkeys by adult Alphitobius diaperinus (Coleoptera: Tenebrionidae). J. Med. Entomol. 37: 480-483.
Cristhiane RohdeI; Luis F.A. AlvesI, V; Pedro M.O.J. NevesII, V; Sérgio B. AlvesIII, V; Everton R.L. da SilvaIV; José E.M. de AlmeidaVI
sebalves[arroba]esalq.usp.br
ILab. Zoologia, CCBS, Unioeste, Campus de Cascavel, R. Universitária, 2069, 85819-110, Cascavel, PR
IIDepto. Agronomia, UEL, C. postal 6001, 86051-990, Londrina, PR
IIIDepto. Entomologia, Fitopatologia e Zoologia Agrícola, ESALQ/USP, C. postal 9, 13418-900, Piracicaba, SP
IVUnioeste, Campus de Marechal Cândido Rondon, CCA, Programa de Mestrado em Agronomia
VBolsista de Produtividade em Pesquisa/CNPq
VILab. Controle Biológico, Instituto Biológico, C. postal 70, 13001-970, Campinas, SP.
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




