 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
O período de desenvolvimento variou de 10,1 a 9,6 dias, diferindo estatisticamente entre as linhagens estudadas. Os menores valores para o período de desenvolvimento foram constatados nas linhagens Lsa4, Lsg1, Lsg11 e Lsa8, porém só detectando-se diferenças significativas entre estas e as linhagens Lsg12 e Lsg5. Observaram-se valores acima de 50% na porcentagem de emergência para a maioria das linhagens estudadas, sendo o maior deles para Lsg1, que diferiu, significativamente, das linhagens, Lsa3, Lsg5, Lsg16, Lsg20, e Lsg23 (Tabela 1). Essa maior porcentagem de emergência observada em ovos de S. frugiperda foi também constatada em diferentes hospedeiros por Sá e Parra (1994), Cônsoli e Parra (1996) e por Greenberg et al. (1998), para T. pretiosum sobre ovos de Helicoverpa zea (Bodd.) (95,0%), Heliothis virescens Fabr.(94,3%) e S. exigua (75,4%), respectivamente. Os períodos de desenvolvimento, embora tenham diferido entre as linhagens, foram próximos aos valores observados por Greenberg et al. (1998) para T. pretiosum mantidos em ovos de H. zea e S. exigua que foram de 9,3 e 9,4 dias, respectivamente, e inferiores aos observados por Sá e Parra (1994) para duas linhagens de T. pretiosum sobre ovos de H. zea: 11,9 e 11,4 dias.
A longevidade de T. pretiosum variou em função da linhagem estudada, sendo que Lsg7, Lsg3, Lsg1, Lsg18, Lsg12, Lsg29 e Lsg11 apresentaram maiores valores em relação às demais, porém só diferiram, significativamente, da linhagens Lsg8, Lg4, Lsg17, Lsg20, Lsa8, Lsa3, Lsa2, Lsg23 e Lsg5 (Tabela 1). As linhagens Lsg18, Lsg11 e Lsg1 tiveram, ainda, maior capacidade de parasitismo (número médio de ovos parasitados e porcentagem de parasitismo) não diferindo significativamente entre si, embora tal parasitismo tenha resultado em menor número de adultos emergidos por ovo de S. frugiperda (Tabela 1). O menor número de indivíduos por ovo do hospedeiro pode ser vantajoso para o desenvolvimento de Trichogramma, já que mais nutrientes estarão disponíveis para a sua assimilação e desenvolvimento, gerando, com isso, indivíduos maiores e mais competitivos. Por outro lado, o aumento no número de adultos por hospedeiro, além de poder diminuir a qualidade do indivíduo gerado, poderá refletir na eficiência de controle, já que poderá resultar numa menor quantidade de ovos parasitados.
Valores distintos quanto a essas características também foram encontrados por Gomes e Parra (1998) para diferentes linhagens de T. pretiosum sobre ovos de A. kuehniella. Segundo esses autores, o parasitismo de T. pretiosum, proveniente de A. kuehniella sobre ovos do hospedeiro natural Anticarsia gemmatalis Hüb. foi baixo, embora o número de adultos emergidos por ovo do hospedeiro tenha sido maior do que o observado no presente estudo. No entanto, Greenberg et al. (1998) observaram maior número de ovos de S. exigua parasitados por T.pretiosum (em média 44,8 ovos por fêmea), porém esses autores forneceram um maior número de ovos, cerca de 400 ovos, o que pode ter contribuído para o aumento na razão de parasitismo.
Tabela 1. Características biológicas (Média ± Erro padrão) de 20 linhagens de T. pretiosum mantidas em ovos isolados de S. frugiperda. 25,0 ± 2,0ºC, U.R. 70 ± 10% e fotofase de 14 horas
|
Duração (dias) (médiaア EP)1, 2 |
Emergência (%) médiaEP) 1 |
Longevidade (dias) médiaア EP) 1, 2 |
Nºde adultos/ovo hospedeiro médiaア EP) 1, |
Nº de ovos parasitados(médiaア EP) 1, 3 |
Parasitismo (%) (médiaア EP) 1, 4 |
Razão sexual (médiaア EP) 1, |
|
|
Ls a2 |
9,7 ア 0,01 bcd |
86,0 ア 3,29 abcd |
3,1 ア 0,30 hi |
1,3 ア 0,04 bcde |
17,03 ア 1,26 ij |
41,5 ア 3,17 ij |
0,80 ア 0,01 ab |
|
Ls a3 |
9,8 ア 0,05 abcd |
55,7 ア 8,92 f |
3,5 ア 0,31 ghi |
1,1 ア 0,05 def |
11,0 ア 1,97 j |
27,5 ア 4,94 j |
0,79 ア 0,05 ab |
|
Ls a8 |
9,7 ア 0,15 cd |
90,91 ア 1,50 abc |
3,8 ア 0,47 ghi |
1,2 ア 0,02 cdef |
24,7 ア 1,35 defgh |
60,0 ア 3,23 efghi |
0,81 ア 0,01 ab |
|
Lsg1 |
9,6 ア 0,05 d |
95,3 ア 0,97 a |
8,9 ア 0,71 ab |
1,2 ア 0,02 cdef |
32,7 ア 0,72 abc |
81,9 ア 1,80 abc |
0,76 ア 0,02 ab |
|
Lsg2 |
9,9 ア 0,02 abcd |
83,0 ア 2,56 abcd |
5,5 ア 0,43 cdef |
1,5 ア 0,04 a |
24,3 ア 1,22 defgh |
60,6 ア 3,08 efgh |
0,75 ア 0,02 ab |
|
Lsg3 |
9,8 ア 0,05 abcd |
82,0 ア 4,63 abcd |
9,0 ア 0,50 a |
1,1 ア 0,03 ef |
25,9 ア 1,27 defg |
64,8 ア 3,19 defg |
0,71 ア 0,03 b |
|
Lsg4 |
9,61 ア 0,07 d |
92,6 ア 1,70 ab |
4,1 ア 0,41 fghi |
1,3 ア 0,05 abcd |
17,0 ア 1,27 ij |
42,6 ア 3,19 ij |
0,71 ア 0,71 b |
|
Lsg5 |
10,1 ア 0,02 ab |
76,0 ア 5,00 bcd |
2,6 ア 0,19 i |
1,4 ア 0,08 ab |
16,1 ア 1,14 j |
39,9 ア 3,17 ij |
0,79 ア 0,02 ab |
|
Lsg7 |
9,7 ア 0,02 abcd |
94,3 ア 0,96 ab |
9,9 ア 0,83 a |
1,1 ア 0,02 f |
27,2 ア 1,69 bcdef |
68,1 ア 4,24 cdef |
0,80 ア 0,02 ab |
|
Lsg8 |
10,0 ア 0,02 abcd |
85,5 ア 4,70 abcd |
4,6 ア 0,37 efgh |
1,2 ア 0,03 bcdef |
23,8 ア 1,84 fghi |
59,6 ア 4,60 efghi |
0,84 ア 0,02 a |
|
Lsg11 |
9,65 ア 0,04 cd |
93,8 ア 1,30 ab |
7,1 ア 0,55 abcd |
1,2 ア 0,02 cdef |
32,8 ア 0,77 ab |
82,0 ア 1,29 ab |
0,72 ア 0,02 b |
|
Lsg12 |
10,1 ア 0,02 a |
88,8 ア 3,43 abc |
7,8 ア 0,57 abc |
1,2 ア 0,02 cdef |
28,9 ア 1,36 abcd |
72,3 ア 3,42 abcd |
0,80 ア 0,02 ab |
|
Lsg16 |
10,0 ア 0,01 abc |
55,8 ア 6,82 ef |
5,3 ア 0,52 defg |
1,4 ア 0,05 ab |
19,7 ア 1,87 ghij |
49,4 ア 4,69 ghij |
0,81 ア 0,01 ab |
|
Lsg17 |
9,7 ア 0,03 abcd |
84,0 ア 1,88 abcd |
4,0 ア 0,59 ghi |
1,3ア 0,04 abc |
27,8 ア 1,21 bcdef |
69,4 ア 3,03 bcdef |
0,74 ア 0,03 b |
|
Lsg18 |
9,8 ア 0,01 abcd |
93,7 ア 0,81 ab |
8,2 ア 0,71 abc |
1,2 ア 0,02 cdef |
34,1 ア 0,65 a |
89,3 ア 1,63 a |
0,79 ア 0,01 ab |
|
Lsg19 |
10,0 ア 0,02 abcd |
90,8 ア 3,56 abc |
6,2 ア 0,63 cde |
1,1 ア 0,02 ef |
26,8 ア 1,73 cdef |
67,4 ア 4,70 def |
0,78 ア 0,02 ab |
|
Lsg20 |
10,0 ア 0,03 abcd |
69,2 ア 5,03 def |
3,8 ア 0,44 ghi |
1,3 ア 0,04 abcd |
19,7 ア 1,55 hij |
49,4 ア 4,16 hij |
0,76 ア 0,03 ab |
|
Lsg23 |
9,9 ア 0,01 abcd |
73,7 ア 4,08 cde |
3,1 ア 0,40 hi |
1,4 ア 0,04 ab |
23,8 ア 1,08 fghi |
59,6 ア 2,73 efghij |
0,75 ア 0,01 ab |
|
Lsg26 |
10,0 ア 0,02 abcd |
86,9 ア 4,70 abcd |
6,43 ア 0,68 bcde |
1,2 ア 0,04 cdef |
25,8 ア 1,75 defgh |
64,6 ア 4,37 defgh |
0,76 ア 0,02 ab |
|
Lsg29 |
9,7 ア 0,02 dcb |
83,5 ア 4,55 abcd |
7,8 ア 0,43 abc |
1,2 ア 0,02 def |
28,4 ア 2,11 bcde |
71,0 ア 5,28 bcde |
0,76 ア 0,02 ab |
|
1Médias seguidas de mesmas letras, não diferem entre si, pelo teste de Tukey (p 0,05); 2, 3, 4Dados transformados em log (X), X2 e X3, respectivamente |
|||||||
A longevidade aliada à capacidade de parasitismo é uma característica importante quando se pretende utilizar um inimigo natural para o controle de uma praga, já que insetos mais longevos permanecem mais tempo no ambiente e são, teoricamente, capazes de parasitar maior número de hospedeiros durante a fase adulta (Gomes e Parra, 1998). Deve-se ressaltar, porém, que a elevada capacidade de parasitismo observada nesta pesquisa foi obtida pela forma individual em que foram oferecidos os ovos, eliminando-se, assim, as barreiras físicas da presença de escamas e distribuição dos ovos em camadas, comuns nas posturas de S. frugiperda.
Os valores da razão sexual, apesar de diferirem entre as linhagens, podem ser considerados elevados (Tabela 1), sendo superiores àqueles observados por Toonders e Sánchez (1987), de 0,51 para Trichogramma sp. em ovos de S. frugiperda e por Sá e Parra (1994), para duas linhagens de T. pretiosum sobre ovos de H. zea, que foram de 0,56 e 0,25. Em geral, os valores encontrados para os parâmetros biológicos avaliados evidenciam a boa adequação dos ovos de S. frugiperda ao desenvolvimento de T.pretiosum, o que sugere a possibilidade de sua utilização para o controle dessa praga.
De acordo com a análise de agrupamento, as linhagens diferiram quanto às características biológicas avaliadas, formando-se grupos distintos segundo o grau de similaridade apresentado por cada grupo (Figura 1). Tal resultado mostra que, apesar dessas linhagens serem provenientes de uma mesma região e de um mesmo hospedeiro, porém coletadas em épocas diferentes, existe uma grande variação nas populações de T. pretiosum, provavelmente associada a uma origem parental diferente desses indivíduos, que resultam em diferenças genotípicas que refletem na capacidade do parasitóide em aceitar e se desenvolver em ovos de S. frugiperda.
Outros fatores podem ter determinado as diferenças entre as linhagens, como as variações que ocorrem no comportamento de seleção hospedeira entre linhagens ou espécies de Trichogramma (Pavlik, 1993), diferenças quanto à própria adequação aos ovos do hospedeiro ou pelo fato de as linhagens utilizadas terem sido mantidas por várias gerações em ovos do hospedeiro alternativo A. kuehniella, dificultando, assim, a aceitação de algumas linhagens por ovos de S. frugiperda. Segundo Li et al. (1994), Trichogramma sp. nr. sibericum Sorok. após ser criado por várias gerações em ovos do hospedeiro alternativo, Peridroma saucia (Hübn.) diminuiu a sua capacidade de aceitar e parasitar ovos de seu hospedeiro natural Rhopobota naevana (Hübn.). Esse fato, entretanto, não ocorreu no presente trabalho, pois todas as linhagens foram mantidas por uma geração em ovos de S. frugiperda antes do início da fase experimental, como forma de minimizar o efeito da criação no hospedeiro alternativo. Assim, as linhagens Lsg1, Lsg11 e Lsg18, foram selecionadas para se testar a preferência hospedeira entre ovos de A.kuehniella e S. frugiperda, por terem apresentado maior capacidade para parasitar e se desenvolver em ovos de S. frugiperda (Tabela 1), além do elevado grau de similaridade (>99,8%) quanto às características biológicas avaliadas (Figura 1).
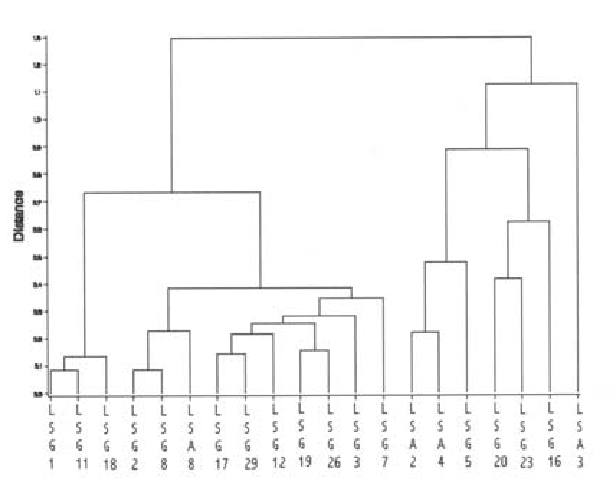
Figura 1. Fenograma resultante da análise de agrupamento de características biológicas de 20 linhagens de T. pretiosum desenvolvidas sobre ovos de S. frugiperda.
A freqüência de contatos entre ovos de S.frugiperda e A. kuehniella foi diferente e altamente significativa, ocorrendo diferenças na preferência hospedeira entre as três linhagens estudadas (Lsg1, Lsg11 e Lsg18). Em geral, T. pretiosum preferiu S. frugiperda a A. kuehniella (Figura 2). As linhagens Lsg11 e Lsg18 tiveram maior freqüência de contatos com ovos de S. frugiperda em relação a ovos de A.kuehniella, se comparadas à linhagem Lsg1, o que revela a preferência daquelas duas por ovos do hospedeiro natural, já que existe uma forte relação entre número de contatos e parasitismo (Wührer e Hassan, 1993). A freqüência de contatos entre ovos do hospedeiro natural e do alternativo foi semelhante para Lsg1 (Figura 2), o que segundo Hassan (1989) pode, com a continuidade da criação em A. kuehniella, levar a uma mudança de preferência em direção ao hospedeiro alternativo e perda de efetividade no campo.
A preferência por ovos de S. frugiperda, das linhagens Lsg11 e Lsg18, mostra que, apesar dessas serem mantidas por várias gerações em ovos de A. kuehniella, não perderam a sua capacidade de encontrar e parasitar ovos do hospedeiro natural, mantendo dessa forma, a sua agressividade.
Entretanto, este resultado não é definitivo, pois nem sempre o maior tempo de contato (exame), significa maior parasitismo. Assim, Wührer e Hassan (1993) observaram que T. pretiosum, apesar de ter contactado ovos de Plutella xylostella (L.) por mais tempo, não apresentou taxas de parasitismo superiores àquelas do hospedeiro alternativo S. cerealella, podendo tal fato, ser atribuído a um maior gasto de tempo no exame dos ovos de P. xylostella.
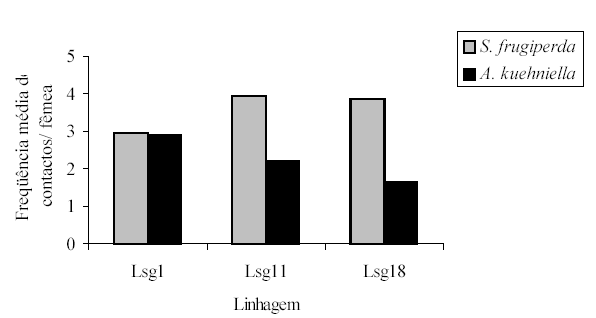
Figura 2. Freqüência média de contactos de fêmeas de três linhagens de T. pretiosum sobre ovos de S. frugiperda e A. kuehniella (÷2= 9,79, P= 0,007). Temperatura de 25,0 ± 2,0 e U.R. 70 ± 10%
No presente trabalho, portanto, verificou-se a existência de variações no comportamento das linhagens com relação a ovos individualizados de S.frugiperda, e que algumas delas não alteram o seu comportamento quando criadas no hospedeiro alternativo, continuando a preferir ovos do hospedeiro natural, como no caso das linhagens Lsg11 e Lsg18.
CÔNSOLI, F.L.; PARRA, J.R.P. Biology of Trichogramma galloi and T. pretiosum (Hymenoptera: Trichogrammatidae) reared in vintro and in vivo. Ann. Entomol. Soc. Am., Lanhan, v. 89, p. 828-834, 1996.
COBERT, S.A. Insect chemosensory responses: a chemical legacy hypothesis. Ecol. Entomol., Blackwell, v.10, p. 143-153, 1985.
CRUZ, I. et al. Efeito do nível de saturação de alumínio em solo ácido sobre os danos de Spodoptera frugiperda (J.E. Smith) em milho. An. Soc. Entomol. Bras., Londrina, v. 25, p.293-297, 1996.
GOMES, S.M.; PARRA, J.R.P. The parasitism as a tool for factitious host selection for Trichogramma galloi Zucchi and T. pretiosum Riley. Mitt. Biol. Bundesanstalt., Berlin, p. 13-23. 1998.
GREENBERG, S.M. et al. A. Parasitism of beet armyworm by T. pretiosum and T. minutum under laboratory and field conditions. Southwest. Entomol., Dallas, v. 23, n. 2, p. 183-188, 1998.
HASSAN, S.A. Selection of suitable Trichogramma strains to control the codling moth Cydia pomonella and the two summer fruit tortrix moths Adoxophyes orana, Pnademis heparana (Lep.: Tortricidae). Entomophaga, Montpellier, v. 34, p. 19-27, 1989.
HASSAN, S.A. Strategies to select Trichogramma species for use in biological control. In: E. Wanjnberg & S. A Hassan (Ed.). Biological control with egg parasitoids. Wallingford: CAB 1994. p. 55-72.
HASSAN, S.A. et al. Der Einfluss des wirtes in der massenzucht auf die qualitat des eiparasiten Trichogramma evanescens bei der bekämpfung des maízünslers, Ostrinia nubilalis. Entomophaga, Montpellier, v. 23, n.4, p. 321-329, 1978.
LI, S. Y. et al. Selection of suitable Trichogramma species for potencial control of the blackheaded fireworm infesting cranberries. Biol. Control., Orlando, v. 4, p. 244-248, 1994.
PARRA, J.R.P. et al. Número ideal de ovos de Anagasta kuehniella (Zeller, 1879) por caixa de criação para pesquisas com Trichogramma spp. An. Soc. Entomol. Bras., Londrina, v. 18, p. 391-402, 1989.
PAVLIK, J. Variability in the host acceptance of european corn borer, Ostrinia nubilalis Hbn. (Lep., Pyralidae) in strains of the egg parasitoid Trichogramma spp. (Hym., Trichogrammatidae). J. Appl. Entomol., Berlin, v. 115, p. 77-84, 1993.
PINTUREAU, B. et al. Etude de quelques facteurs de variation de la fécondité chez Trichgoramma maidis Pintureau & Voegele (Hym.: Trichogrammatidae). Agronomiae, Paris, v.1, n. 4, p. 315-322, 1981.
ROA, F. G. Effectiveness of releases of Trichogramma spp. in crops Colômbia. Trichogramma News, Braunschweig, v. 8, p. 30, 1995.
SÁ, L.A.; PARRA, J.R.P. Biology and parasitism of Trichogramma pretiosum Riley (Hym., Trichogrammatidae) on Ephestia kuehniella (Zeller) (Lep., Pyralidae) and Heliothis zea (Boddie) (Lep., Noctuidae) eggs. J. Appl. Entomol., Berlin, v. 118, p. 38- 43, 1994.
TOONDERS, T.J.; SÁNCHEZ, J.L.C. Evaluation de la efectividad de Trichogramma spp. (Hymenoptera: Trichogrammatidae) en el combate de Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae). recomendaciones para su uso. Cent. Entomol. Acar., Chapingo, p.75-84, 1987.
WÜHRER, B.G.; HASSAN, S.A. Selection of effective species/strains of Trichogramma (Hym., Trichogrammatidae) to control the diamondback moth Plutella xylostella L. (lep., Plutellidae). J. Appl. Entomol., Berlin, v. 116:, p. 80-89, 1993.
Received on January 08, 2003.
Accepted on September 22, 2003.
Eduardo Barbosa Beserra1*, Carlos Tadeu dos Santos Dias2 e José Roberto Postali Parra3
ctsdias[arroba]carpa.ciagri.usp.br
1. Departamento de Farmácia e Biologia, Universidade Estadual da Paraíba, João Pessoa, Paraíba, Brasil.
2. Departamento de Ciências Exatas, Esalq, Universidade de São Paulo.
3. Departamento de Entomologia, Fitopatologia e Zoologia Agrícola, Esalq/ Universidade de São Paulo. *Autor para correspondência.
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




