 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
O experimento foi realizado no Departamento de Produção Vegetal da USP/ESALQ, em Piracicaba, SP, Brasil. Foram utilizados mamões 'Sunrise Solo' linhagem 72/12, colhidos no município de Teixeira de Freitas, BA. A colheita foi realizada no dia 24/05/2000 e os frutos foram transportados até a CEAGESP, na cidade de São Paulo, SP, onde chegaram no dia 26/05/2000.
No recebimento, os mamões foram imediatamente selecionados. Utilizaram-se frutos sem defeitos, com peso médio de 400 ± 27 g, em dois estádios de maturação: a) verde: frutos com coloração verde e uma lista amarela e, b) maduro: frutos com coloração verde e três ou quatro listas amarelas.
Os frutos foram submetidos a 0, 30, 90 e 270 nL L-1 de 1-MCP. A aplicação constou da colocação dos frutos em câmara hermética e exposição ao produto por um período de 12 horas sob temperatura de 20oC. Os frascos foram abertos no interior das câmaras, as quais foram fechadas imediatamente para evitar a perda do gás. Após 12 horas de tratamento, as câmaras foram abertas e os frutos foram colocados em condições ambientais (20°C), sendo avaliados a cada dois dias, durante oito dias.
O delineamento experimental adotado foi inteiramente casualizado em esquema fatorial para cada estádio de maturação. Foram utilizadas quatro repetições de cinco frutos por parcela.
As variáveis determinadas foram: a) coloração da casca: usando-se colorímetro, com os resultados expressos em ângulo de cor (h°). O h° define a coloração básica, onde 0° = vermelho, 90° = amarelo e 180° = verde (McGuire, 1992); b) firmeza de polpa: pelo método da aplanação (Calbo & Nery, 1995), c) podridões: foi calculada a porcentagem de frutos afetados em cada repetição e a severidade das lesões (frutos não afetados, pouco ou muito afetados). Foram considerados pouco afetados aqueles frutos com início de podridão na região peduncular e/ou lesão £ 1 cm na superfície da casca. O agente causal foi identificado por sintomatologia e análise microscópica; d) teor de sólidos solúveis: em refratômetro digital; e) taxa respiratória e f) produção de etileno.
Para a determinação da taxa respiratória e da produção de etileno, amostras de frutos foram acondicionadas em câmaras herméticas, que foram mantidas fechadas durante três horas. Posteriormente, coletou-se uma amostra de 2 L em um recipiente de polietileno tereftalato que foi enviado ao laboratório do Núcleo de Pesquisa em Pós-Colheita da UFSM, em Santa Maria, RS. Determinou-se a concentração de etileno em cromatógrafo a gás com coluna Porapak N e detector de ionização de chama (FID). A temperatura da coluna, injetor e detector foi de 90, 140 e 200°C, respectivamente. De cada amostra de gás, foram extraídas duas amostra de 1 mL com seringa plástica descartável e injetadas no cromatógrafo. As áreas de pico de etileno foram transformadas em ppm através de um fator de correção obtido pela injeção de um gás padrão com 10 ppm de etileno. Para o cálculo da produção de etileno, foram levados em consideração o volume da câmara de armazenamento (60 L), a massa dos frutos e o tempo de acúmulo de etileno.
Para a determinação da respiração (produção de CO2) utilizou-se um analisador de gases (CO2 e O2). Os gases das amostras foram conduzidos pelo analisador até a estabilização dos valores das concentrações de CO2, expressos em porcentagem. Também para o cálculo da respiração, foram levados em consideração o volume da câmara de armazenamento, a massa de frutos e o tempo de acúmulo de CO2.
Os resultados foram submetidos a análise de variância, comparação de médias e regressão polinomial.
Os frutos do estádio verde tratados com 90 ou 270 nL L-1 de 1-MCP apresentaram perda de coloração verde mais lenta do que os frutos não tratados ou tratados com 30 nL L-1 (Tabela 1). Para os frutos do estádio maduro verificou-se diferença consistente apenas entre os frutos tratados com 270 nL L-1 e os frutos não tratados. O desverdecimento é decorrente da quebra da estrutura da molécula de clorofila, envolvendo a atividade da enzima clorofilase. A elevação da atividade desta enzima está geralmente associada com a produção de etileno durante o amadurecimento do fruto (Tucker, 1993). O 1-MCP se liga ao sítio de ligação do etileno na célula evitando a ação do mesmo sobre os processos fisiológicos de amadurecimento (Serek et al., 1995). Assim, a perda de cor verde, resultante do processo normal de amadurecimento, foi retardada com a aplicação do 1-MCP. A retenção do desenvolvimento da coloração da casca em frutos tratados com 1-MCP também foi verificada em ameixa (Abdi et al., 1998), banana (Golding et al., 1998), tomate (Dupille & Sisler, 1995) e abacate (Fan et al., 2000).
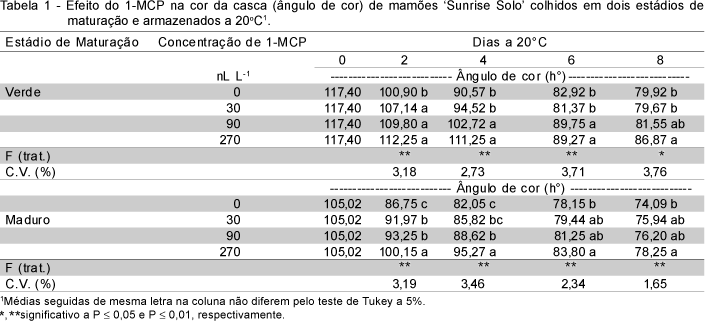
O 1-MCP afetou significativamente a porcentagem de frutos com podridões ao longo do período de armazenamento (Tabela 2). Os fungos mais frequentemente encontrados foram os dos gêneros Colletrotrichum e Phomopsis.

Para o estádio verde, as podridões aumentaram sensivelmente aos 6 dias de conservação a 20oC. Os frutos do controle ou tratados com 30 nL L-1 de 1-MCP apresentaram porcentagem de podridão significa-tivamente superior aos frutos tratados com 90 ou 270 nL L-1. Embora neste período tenha sido observada porcentagem de 15 a 20% de podridão nos frutos tratados com as maiores concentrações, estas podridões foram consideradas de baixa severidade (Tabela 3), tendo sido constatado apenas início de podridão peduncular, sendo a área afetada inferior a 5% da superfície do fruto. Assim, aos 6 dias de conservação a 20oC, os frutos tratados com 90 ou 270 nL L-1 de 1-MCP encontravam-se aptos para a comercialização, ao contrário dos frutos controle ou tratados com 30 nL L-1. Aos 8 dias, todos os tratamentos deste estádio encontravam-se com alta porcentagem de podridão e alta severidade das lesões (Tabelas 2 e 3).

Para o estádio maduro, os frutos não apresentaram podridão aos 2 dias de armazenamento a 20oC, ao passo que aos 4 dias a porcentagem de podridão variou de 15 a 20% (Tabela 2). Os frutos não tratados ou tratados com 30 nL L-1 apresentaram maior severidade de podridão (Tabela 3). Nesses, 50% apresentaram podridão peduncular e lesões com mais de 1cm de diâmetro na casca, o que descartou os frutos deste tratamento para a comercialização aos 4 dias. Os frutos tratados com 90 ou 270 nL L-1 de 1-MCP apresentaram, neste período, percentual de podridão de 15%, sendo que a totalidade dos frutos afetados apresentaram início de podridão peduncular e ausência de lesões na casca. Aos 6 dias de conservação todos os tratamentos estavam inaptos para a comercialização, considerando as altas porcentagens de podridão e a severidade das lesões.
Os frutos tratados com 1-MCP, principalmente nas concentrações de 90 nL L-1 e 270 nL L-1 apresentaram menor perda de firmeza durante os períodos de avaliação (Tabela 4), diferindo significativamente do controle em ambos estádios de maturação. Frutos tratados com 30 nL L-1 de 1-MCP apresentaram comportamento intermediário. A firmeza de polpa do fruto é determinada pela força de coesão entre as pectinas. Com a evolução do amadurecimento ocorre atuação de enzimas pectinolíticas, que transformam a pectina insolúvel em solúvel e promovem o amolecimento dos frutos. O amolecimento dos frutos é um dos processos do amadurecimento mais sensíveis ao etileno (Lelièvre et al., 1997a) . A maior firmeza dos frutos tratados com as maiores concentrações de 1-MCP está provavelmente associada à redução da atividade das enzimas pectinolíticas, causada pela redução da ação do etileno. Resultados similares a estes foram obtidos em maçã (Fan et al., 1999) e banana (Jiang et al., 1999).

O teor de sólidos solúveis não foi influenciado pelo 1-MCP, em ambos estádio de maturação; variou de 11,15 a 12,01oBrix para o estádio verde e de 12,74 a 13,62oBrix para o estádio maduro, durante o amadurecimento.
O 1-MCP reduziu a taxa respiratória e a produção de etileno dos frutos nos dois estádios de maturação (Figura 1). As concentrações de 90 e 270 nL L-1 apresentaram as maiores reduções nestes processos, enquanto que, na concentração de 30 nL L-1, os resultados pouco diferiram do controle. O aumento na produção de etileno precedeu a ascensão climatérica da respiração, mostrando o efeito indutor do etileno sobre o processo respiratório.

O 1-MCP é caracterizado por ser um competidor pelo sítio de ligação do etileno na célula. Quando aplicado no momento correto, o 1-MCP ocupa os sítios de ligação do etileno e impede os seus efeitos, como por exemplo a síntese de enzimas degradativas, aumento na taxa respiratória e a própria produção de etileno (autocatálise). A capacidade de conservação de um produto hortícola está inversamente relacionada à taxa respiratória, e em muitos casos, com a taxa de produção de etileno (Kader, 1994). A redução na produção de etileno e na taxa respiratória explicam a maior conservação dos frutos tratados com 90 e 270 nL L-1 de 1-MCP. A menor taxa respiratória e menor produção de etileno de frutos tratados com 1-MCP também foi reportada em trabalhos anteriores (Abdi et al., 1998; Golding et al., 1998; Fan et al., 1999; Jiang et al., 1999).
A firmeza da polpa e a porcentagem de podridão foram considerados os principais fatores que influenciaram a vida de prateleira dos frutos deste experimento. Desta forma, para frutos no estádio verde tratados com 1-MCP (90 ou 270 nL L-1), a máxima capacidade de armazenamento foi de 6 dias, enquanto que os frutos não tratados ou os tratados com 30 nL L-1 apresentaram vida de prateleira de 4 dias. Para o estádio maduro, a máxima capacidade de armazenamento foi de 4 dias para os frutos tratados com 1-MCP (90 ou 270 nL L-1) e 2 dias para os não tratados ou tratados com 30 nL L-1.
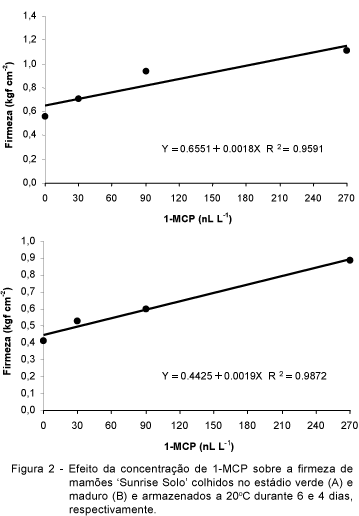
Levando-se em consideração a firmeza de polpa realizou-se regressão polinomial para as concentrações de 1-MCP nos máximos períodos de armazenamento para os dois estádio de maturação. Para tal, fixou-se o tempo máximo de conservação observado no presente experimento em seis dias para o estádio verde e quatro dias para o estádio maduro. Tanto no estádio verde (Figura 2A) quanto no estádio maduro (Figura 2B) esta variável apresentou comportamento linear. Portanto, para determinar a máxima capacidade de manutenção de firmeza seria necessário avaliar concentrações superiores a 270 nL L-1. Por outro lado, frutos com firmeza acima de 1,0 kgf cm-2 encontravam-se excessivamente firmes. Isso pode reduzir a aceitação comercial dos frutos, pois, embora a coloração externa estivesse amarela, os frutos não amoleceram adequadamente. Este comportamento foi observado nos frutos mais verdes (ausência de listas amarelas) tratados com 270 nL L-1 de 1-MCP.
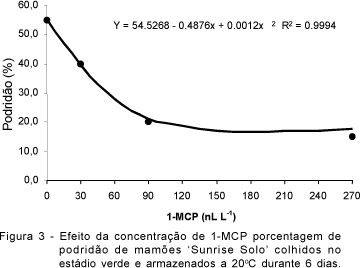
Para a porcentagem de podridão, as concentrações de 90 e 270 nL L-1 de 1-MCP promoveram os melhores resultados, com valores entre 15 e 20% (Figura 3).
Para o estádio maduro não houve significância das concentrações de 1-MCP sobre o percentual de podridões aos quatro dias (Tabela 2), enquanto que para a variável firmeza verificou-se significância da regressão linear, ou seja, a firmeza aumenta à medida em que a concentração de 1-MCP é incrementada (Figura 2).
A aplicação do 1-MCP aumenta a vida de prateleira de mamões, como consequência da capacidade do 1-MCP de inibir a ação do etileno nos tecidos e retardar o amadurecimento. O aumento no período de conservação sob condições ambientais é importante, considerando a alta perecibilidade do mamão após a colheita. O ganho de dois a três dias de vida de prateleira pode proporcionar transporte dos frutos a maiores distâncias e ampliação do período de comercialização dos mesmos.
ABDI, N.; McGLASSON, W.B.; HOLFORD, P.; WILLIAMS, M; MIZRAHI, Y. Responses of climateric and suppressed-climacteric plums to treatment with propylene and 1-methylcyclopropene. Postharvest Biology and Technology, v.14, p.29-39, 1998.
BLANKENSHIP, S.M.; SISLER, E.C. 2,5-norbornadiene retards apple softening. HortScience, v.24, p.313-314, 1989.
BLANKENSHIP, S.M.; SISLER, E.C. Response of apples to diazocyclopentadiene inhibition of ethylene binding. Postharvest Biology and Technology, v.3, p.95-101, 1993.
BURG, S.P.; BURG, E.A. Molecular requirements for the biological activity of ethylene. Plant Physiology, v.42, p.144-152, 1967.
CALBO, A.G.; NERY, A.A. Medida de firmeza em hortaliças pela técnica de aplanação. Horticultura Brasileira, v.13, p.14-18, 1995.
DUPILLE, E.; SISLER, E.C. Effects of ethylene receptor antagonist on plant material. In: AIT-OUBAHOU, A.; EL-OTMANI, M. (Ed.) Postharvest, pathology and technologies for horticultural commodities: recents advances. Agadir: Institute Agronomique et Veterinaire Hassan II., 1995. p.294-301.
FAN, X.; ARGENTA, L.; MATTHEIS, J.P. Inhibition of ethylene action by 1-methylcyclopropene prolongs storage life of apricots. Portharvest Biology and Technology, v.20, p.135-142, 2000.
FAN, X.; BLANKENSHIP, S.M.; MATTHEIS, J.P. 1-methylcyclopropene inhibits apple ripening. Journal of the American Society for Horticultural Science, v.124, p.690-695, 1999.
FENG, X.; APELBAUM, A.; SISLER, E.C.; GOREN, R. Control of ethylene responses in avocado fruit with 1-methylcyclopropene. Portharvest Biology and Technology, v.20, p.143-150, 2000.
GOLDING, J.B.; SHEARER, D.; WYLLIE, S.G.; McGLASSON, W.B. Application of 1-MCP and propylene to identify ethylene-dependent ripening processes in mature banana fruit. Portharvest Biology and Technology, v.14, p.87-98, 1998.
GONG, Y.; TIAN, M.S. Inhibitory effect of diazocyclopentadiene on the development of superficial scald in 'Granny Smith' apples. Plant Growth Regulation, v.26, p.117-121, 1998.
JIANG, Y.; JOYCE, D.C.; MACNISH, A.J. Extension of the shelf life of banana fruit by 1-methylcyclopropene in combination with polyethylene bags. Postharvest Biology and Technology, v.16, p.187-193, 1999.
KADER, A.A. Postharvest technology of horticultural crops. Oakland: University of California, 1994. 296p.
LELIÈVRE, J.M.; TICHIT, L.; DAO, P.; FILLION, L.; NAM, Y.W.; PECH, J.C.; LATCHÉ, A. Effects of chilling on the expression of ethylene biosynthetic genes in Passe-Crassane pear (Pyrus communis L.) fruits. Plant Molecular Biology, v.33, p.847-855, 1997b. www.bireme.br
LELIÈVRE, J.M.; LATCHÉ, A.; JONES, B.; BOUZAYEN, M.; PECH, J.C. Ethylene and fruit ripening. Physiologia Plantarum, v.101, p.727-739, 1997a.
McGUIRE, R.G. Reporting of objective color measurements. HortScience, v.27, p.1254-1255, 1992.
NAKATSUKA, A.; SHIOMI, S.; KUBO, Y.; INABA, A. Expression and internal feedback regulation of ACC synthase and ACC oxidase genes in repening tomato fruit. Plant and Cell Physiology, v.38, p.1130-1110, 1997.
PAULL, R.E. Pineapple and papaya. In: SEYMOUR, G.B.; TAYLOR, J.E.; TUCKER, G.A. (Ed.) Biochemistry of fruit ripening. London: Chapman & Hall, 1993. p.291-323.
PORAT, R.; HALEVY, A.H.; SEREK, M.; BOROCHOV, A. An increase in ethylene sensitivity following pollination is the initial event triggering an increase in ethylene production and enhanced senescence of Phalaenopsis orchid flowers. Physiologia Plantarum, v.88, p.243-250, 1995.
SEREK, M.; SISLER, E.C.; REID, M.S. 1-methylcyclopropene, a novel gaseus inhibitor of ethylene action, improves the life of fruit, cut flowers and potted plants. Acta Horticulturae, v.394, p.337-345, 1995.
SEREK, M.; SISLER, E.C.; REID, M.S. A volatile ethylene inhibitor improves the postharvest life of potted roses. Journal of the American Society for Horticultural Science, v.119, p.572-577, 1994.
SIMÃO, S. Tratado de fruticultura. Piracicaba: FEALQ, 1998. 760p.
SISLER, E.C.; SEREK, M. Inhibitors of ethylene responses in plants at the receptors level: recent developments. Physiologia Plantarum, v.100, p.577-582, 1997.
SISLER, E.C.; SEREK, M.; DUPILLE, E. Comparison of cyclopropene, 1-methylcyclopropene, and 3,3-dimethylcyclopropene as ethylene antagonists in plants. Plant Growth Regulation, v.18, p.169-174, 1996.
TUCKER, G.A. Introduction. In: SEYMOUR, G.B.; TAYLOR, J.E.; TUCKER, G.A. (Ed.) Biochemistry of fruit ripening. London: Chapman & Hall, 1993. p.1-51.
Angelo Pedro Jacomino1,4; Ricardo Alfredo Kluge2,4; Auri Brackmann3,4; Paulo Roberto de Camargo e Castro2 - brackmann[arroba]ccr.ufsm.br
1Depto. de Produção Vegetal - USP/ESALQ, C.P. 9 - CEP: 13418-900 - Piracicaba, SP. 2Depto. de Ciências Biológicas - USP/ESALQ. 3Depto. de Fitotecnia - UFSM - CEP: 97119-900 - Santa Maria, RS. 4Bolsista CNPq.
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




