- Introducción
- Fotosíntesis
- Importancia de la
fotosíntesis - Radiación fotosintética
activa - El
proceso fotosintético - Inhibición de la
fotosíntesis - Conclusión
- Bibliografía

Introducción
El presente trabajo de investigación tiene como
tema principal la fotosíntesis. El proceso de la
fotosíntesis, es un proceso en el cual organismos como
algas y vegetales convierten la energía solar en
energía química, todo esto para posibilitar la
síntesis del carbono. Este proceso permite que organismos
como los vegetales desarrollen infinidad de moléculas
orgánicas a partir de compuestos inorgánicos, de
allí que todos los demás organismos no
autótrofos obtienen las biomoléculas necesarias
para la vida.
El proceso de fotosíntesis es vital para el
crecimiento y desarrollo de una planta, por tanto es importante
comprender las rutas que siguen todos los compuestos que ingresan
al vegetal; como así los factores que favorecen o afectan
a la fotosíntesis, de esa forma podemos comprender mejor
las condiciones necesarias para una producción vegetal
óptima, que es a lo que todo profesional quiere
llegar.
Fotosíntesis
CONCEPTO
La vida en nuestro planeta se mantiene fundamentalmente
gracias a la fotosíntesis que realizan las algas, en el
medio acuático, y las plantas, en el medio terrestre, que
tienen la capacidad de sintetizar materia
orgánica (imprescindible para la constitución
de los seres
vivos) partiendo de la luz y la materia
inorgánica. Se estima que, cada año, los
organismos fotosintetizadores fijan en forma de materia
orgánica en torno a 100.000 millones de toneladas de
carbono.
Etimológicamente fotosíntesis significa:
síntesis con ayuda de la luz. La fotosíntesis es en
esencia, un proceso de óxido reducción, en el que
el carbono del CO 2 se reduce a carbono orgánico. Aunque
en algunos microorganismos fotosintéticos el proceso es
algo diferente, en las plantas consiste básicamente en la
producción de una sustancia orgánica, (un
glúcido sencillo – el adenosín
trifosfato ATP la primera molécula en la que queda
almacenada esa energía química) a partir de
moléculas inorgánicas (el dióxido de carbono
como sustrato a reducir y el agua como dador de electrones que se
oxida) mediante el aprovechamiento de la energía
lumínica (que queda almacenada como energía
química dentro de la molécula sintetizada) y con
desprendimiento de CO2. Con posterioridad, el ATP se usa para
sintetizar moléculas orgánicas de mayor
estabilidad.
La fotosíntesis se puede representar con la
formula mínima de los monosacáridos mediante la
siguiente ecuación:
Luz
CO2 + H2O ? (CH20) + O2
En la fotosíntesis, se libera el oxígeno
integrante de la molécula del agua y se almacena, por
medio de una reacción de reducción, numerosos
compuestos carbonatados que constituyen la materia viva. En
sí, la fotosíntesis es un proceso de oxido –
reducción, en que un donador de electrones, el agua, se
oxida y, un aceptor, el anhídrido carbónico u otro
aceptor adecuado que puede ser el nitrato o sulfato, se reduce.
Esta es una propiedad característica que tienen los
autótrofos, de asimilar CO2 atmosférico y convertir
la energía luminosa en energía
química.
La fotosíntesis es considerada como
un proceso, en el cual se desarrollan tres fases:
1. La absorción de luz y
retención de energía lumínica.
2. La conversión de energía
lumínica en potencial químico.
3. La estabilización y almacenaje de
potencial químico.
IMPORTANCIA DE LA
FOTOSÍNTESIS
Las plantas son indispensables en nuestras vidas por la
fotosíntesis. La fotosíntesis es el
proceso por el cual las plantas y otros determinados organismos
utilizan la energía solar para fabricar sus propios
alimentos, transformando el dióxido de carbono y el agua
en azúcares que almacenan energía química.
Los animales y otros organismos, incapaces de fabricar sus
propios alimentos, únicamente pueden sobrevivir obteniendo
directa o indirectamente de las plantas.
La fotosíntesis sustenta la vida en
la Tierra
La práctica totalidad de la vida en la tierra
depende del agua y de la energía del sol. Sin embargo,
únicamente las plantas, algas y bacterias
fotosintéticas pueden utilizar estos ingredientes de forma
directa para sobrevivir. Con luz solar, dióxido de
carbono, agua y unos pocos minerales del suelo, un vegetal es
capaz de fabricar su propio alimento, pero ningún animal
puede vivir solo a base de estos ingredientes. Casi una cuarta
parte del cerca de millón y medio de especies de
organismos vivos conocidas son fotosintéticas.
Las plantas, bacterias y algas llevan a cabo casi el
total de la fotosíntesis del planeta, lo que les otorga un
lugar preponderante en la Biosfera. Las plantas son la principal
fuente de fotosíntesis en la tierra, en tanto que las
algas, junto con las bacterias fotosintéticas, contribuyen
a la realización de la fotosíntesis en medios
acuáticos.
La fotosíntesis sustenta la vida de
tres formas:
1) Hoy día, los científicos creen que la
fotosíntesis produce casi todo el oxígeno del
mundo. Durante este proceso, los vegetales rompen las
moléculas de agua (H2 O) y producen oxígeno (O2).
La mayoría de los organismos, incluidos los vegetales y
animales, necesitan oxigeno para liberar la energía
almacenada en los alimentos.
2) La mayor parte de los organismos obtiene su
energía directa o indirectamente de la
fotosíntesis. Los animales y la mayoría de los
organismos no fotosintéticos obtienen la energía
alimentándose de vegetales o de otros organismos que han
ingerido vegetales. En este sentido, los vegetales o cualquier
otro organismo fotosintético es el origen de cualquier
cadena alimenticia, una secuencia de transferencia alimenticia
desde un organismo al siguiente que inicia con el organismo que
produce el alimento. Los organismos fotosintéticos se
conocen como productores primarios y se constituyen como la base
de una cadena alimenticia. Los vegetales son los productores
primarios de las terrestres en tanto que las algas y bacterias
fotosintéticas lo son de las acuáticas.
3) Los azúcares producidos por la
fotosíntesis son los bloques de construcción de la
vida. Los vegetales producen azúcares y moléculas
relacionadas mediante la fotosíntesis y los procesos
derivados de ella, y posteriormente combinan estos productos con
minerales del suelo para dar lugar a una amplia variedad de
compuestos. Un vegetal utiliza estos compuestos para determinar
sus características estructurales y fisiológicas.
Al comer vegetales o animales que han comido vegetales, un animal
recibe los compuestos producidos originariamente por la
fotosíntesis, que a su vez emplea para generar su propia
estructura.
RADIACIÓN
FOTOSINTÉTICA ACTIVA.
Apenas 5% de la energía solar que alcanza la
tierra puede ser convertida en carbohidratos a través de
la fotosíntesis foliar debido a que la mayoría de
la fracción de luz incidente es de onda muy corta o muy
larga para ser absorbida por los pigmentos
fotosintéticos.
De la radiación que llega a la tierra, el 44% se
sitúan en la región de las radiaciones visibles que
son utilizadas en la fotosíntesis. La radiación
solar alcanza su máximo en verano (diciembre –
enero) y su mínimo en invierno (junio – julio).
Durante el día, la intensidad máxima de
radiación solar ocurre en la proximidad del
mediodía.
El sol funciona como un inmenso horno, en el cual el
ión hidrógeno se funde produciendo helio y
liberando una cantidad fantástica de energía en la
forma de radiación electromagnética constituido de
diferentes radiaciones, conforme al espectro
electromagnético siguiente.

La luz solar es aparentemente blanca pero es separada en
diferentes colores de luz por el pasaje a través de un
prisma.
La luz se propaga en forma de ondas. Estas ondas
electromagnéticas incluyendo las luminosas tienen una
longitud que es la distancia de un pico de onda a otro. La
diferencia de color entre los rayos luminosos depende realmente
de sus longitudes de onda. La energía que llevan es
inversamente proporcional a la longitud de onda: a menor onda,
mayor energía.


El espectro electromagnético está
constituido por ondas electromagnéticas de distinta
frecuencia, v (o longitud de onda. X = 1/v),
abarcando desde las ondas de mayor longitud y menor
energía, como las ondas de radio y televisión,
hasta las ondas de menor longitud y mayor energía, como
los rayos gamma. La radiación luminosa ocupa una
pequeña franja del espectro, que va desde los 400 a los
700 nm, y se sitúa entre las radiaciones ultravioletas
(UV) y las infrarrojas (IR), y constituye la llamada
radiación fotosintéticamente activa
(PAR).
La radiación luminosa también se llama
radiación visible (VIS) porque el ojo humano es un
órgano sensible foto-detector que nos permite ver esa
radiación. La luz blanca está constituida por la
conjunción de todas las radiaciones luminosas de distinto
color: luz violeta, azul, verde, amarilla, naranja y roja. La luz
blanca se puede descomponer en todo el arco cromático de
luces de color por difracción a través de un prisma
cristalino. Un compuesto o sustancia con color absorbe la luz del
color complementario a la luz del color que muestra, es decir,
ese compuesto es del color de la luz no absorbida, que es la que
vemos reflejada o transmitida.
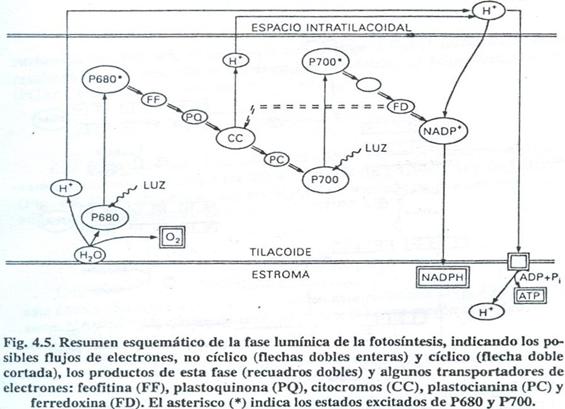
EL PROCESO
FOTOSINTÉTICO.
El proceso fotosintético abarca
numerosos pasos que se agrupan en dos fases: la fase
lumínica y la fase oscura.
FASE LUMÍNICA
Las reacciones de la fase lumínica tienen lugar
en los tilacoides de los cloroplastos donde se sitúan los
pigmentos fotosintéticos y allí, la energía
solar o incluso artificial, es transformada en energía
química depositada en moléculas de ATP y poder
reductor representado por la forma reducida del NADPH
(Nicotinamida –adenín-dinucleótido- fosfato)
sustancia con una fuerte tendencia a reducir otros compuestos.
Ambos compuestos serán empleados en las reacciones que
constituyen la fase oscura, en la que el CO2 será reducido
a carbono orgánico.
Absorción de la luz y transporte
electrónico fotosintético
Para producir energía química a partir de
energía solar, hay que mover electrones. La luz solar
llega en forma de radiaciones electromagnéticas y estas
radiaciones se componen de unidades elementales llamadas fotones
de luz. Cuando una molécula de clorofila absorbe un
fotón, pasa a un estado inestable de mayor energía,
denominado estado excitado, en el que un electrón
periférico se desplaza hacia una posición orbital
superior de un nivel energético mayor. Si este
electrón pasa a otra molécula, la energía se
habrá transmitido y la molécula de clorofila
permanecerá excitada; para volver a su estado fundamental
deberá recibir otro electrón que ocupe el hueco
dejado por el primero. Cuando coexisten numerosas
moléculas de clorofilas agrupadas y ordenadas, la
energía absorbida por cualquiera de ellas puede
transmitirse por resonancia a todo el conjunto, sin que haya
transferencia de electrones. Ambos tipos de transferencia de
energía tienen lugar en el proceso de absorción de
luz por los pigmentos.


En el cloroplasto, los pigmentos están
estrechamente asociados a proteínas y se alojan en la
bicapa lipídica de los tilacoides. Estos complejos
proteína-clorofila forman agrupaciones, cada una con unas
200 – 300 moléculas de clorofila que son las antenas
(reciben la energía lumínica y lo transfiere por
resonancia al centro de reacción), dispuestos alrededor de
un núcleo o centro de reacción, constituido
probablemente por una o dos moléculas de clorofila. Al
recibir la energía, la clorofila del centro de
reacción pierde un electrón que es transferido a
una serie de transportadores de electrones. Los transportadores
actúan en cadena, captando el electrón
(reduciéndose) y seguidamente cediéndolo
(oxidándose) a la siguiente molécula.
También los carotenoides captan energía en sus
longitudes de onda características y lo transfieren a las
clorofilas (también cumplen funciones protectoras al
absorber excesos de energía que podrían formar
substancias nocivas). Esta transferencia de un electrón
sería el paso crucial en la transformación de la
energía luminosa en química. Figura:
absorción de fotones de luz por los pigmentos
clorofílicos
Se distinguen dos tipos de agrupaciones
clorofílicas en las tilacoides, llamadas sistema de
pigmento I y sistema de pigmento II o bien fotosistema I (FSI) y
fotosistema II (FSII), que se diferencian por la proporciones de
clorofila a y b, en las características de sus centros de
reacción y en los transportadores de electrones que los
acompañan. El fotosistema I se localiza en la grana y su
centro de reacción se denomina P700, en tanto que el
fotosistema II está ubicado en las lamelas y en la
periferia de las granas y su centro de reacción se
denomina P680.
La energía luminosa para el
fotosistema I es capturada por la clorofila a (683nm), el
P-
700 (llamado así, debido a que es sensible a
longitudes de onda de 700 nanómetros) y los carotenos. La
energía luminosa destinada al fotosistema II, es capturada
por la clorofila b (673nm), el P-680 (por su particular
sensibilidad de absorber longitudes de onda de
680nm y ficobilinas.

Durante las reacciones de la fase lumínica los
dos fotosistemas actúan coordinadamente, según
veremos a continuación.
La energía absorbida por el FSI provoca la
pérdida de un electrón del P700, que queda en un
estado inestable, con un "hueco" electrónico que
será "rellenado" por un electrón procedente del
FSII. El electrón perdido por el P700 pasa a una cadena de
transportadores que se van reduciendo (al aceptarlo) y oxidando
(al transferirlo) sucesivamente, con un nivel energético
menor en cada paso. Luego de varios compuestos intermedios
aún poco conocidos, el electrón pasa a la
ferredoxina y por último reduce al NADP+ (forma oxidada
del NADPH). La forma reducida se expresa en la ecuación
siguiente:
NADP+ + 2e- + H+ ? NADPH
Se observa que es necesaria la presencia de un
protón (H+) proveniente del espacio intratilacoidal y dos
electrones cedidos por el P700, razón por el cual el flujo
electró nico del FSI deberá tener lugar dos veces
para reducir cada molécula de NADP+.
El FSI se convierte así en un fuerte reductor
capaz de producir NADP+ que será utilizado en la fase
oscura para reducir EL CO2 a carbono orgánico.
Por otra parte, la energía absorbida por el FII
provoca la pérdida de un electrón del P680 que
queda en un estado inestable. El electrón perdido por el
P680 reduce en primer lugar a la feofitina, pasando de
ésta a una serie de transportadores de nivel
energético cada vez menor: plastoquinona, citocromos y
plastocianina. De este último compuesto, el
electrón pasa a ocupar el hueco electrónico del
P700 que de esta manera vuelve a su estado elemental y queda
listo para volver a absorber energía. Ahora, es el P680 es
el que ha quedado con un hueco que será ocupado por un
electrón proveniente de la oxidación del
agua.
El P680 se comporta como un fuerte oxidante que, en su
estado inestable, es capaz de inducir la oxidación del
agua, en la que se desprende O2 conforme a la ecuación
siguiente:
2H2O ? O2 + 4H+ + 4e-
A través de ciertos transportadores, los
electrones liberados pasan a ocupar el hueco electrónico
del P680 queda así listo para volver a absorber
energía. Los protones liberados pasan a acumularse en el
espacio intratilacoidal, de donde proceden los H+ para reducir al
NADP+.

El espacio intratilacoidal se acidifica por la
acumulación de los protones que pierde el agua al oxidarse
y también con protones que se originan durante el
transporte de electrones de la plastoquinona a los citocromos en
el FSI. La concentración de H+ en este compartimento pasa
a ser mucho mayor que en el estroma y se genera así, un
potencial de membrana; los protones tenderán a salir al
estroma a través de canales de membrana específicos
acoplados a la enzima ATPasa, liberando una energía que se
utiliza en la síntesis de ATP a partir de ADP y Pi en un
proceso característico de la fase lumínica llamado
fosforilación no cíclica.

El recorrido de los electrones en el FSI también
puede seguir un camino cíclico, regresando el
electrón del P700 a esta misma molécula (a
través de citocromos y plastocianina); en este caso,
también se verifica un bombeo de protones al espacio
intratilacoidal que permite la síntesis de ATP adicional
(fosforilación cíclica) pero no se generará
poder reductor, ya que los electrones no llegan al NADP +, ni se
liberará oxigeno, porque no podrá haber
oxidación del agua. En la cíclica sólo
actúa el fotosistema I.
Generalmente ambas reacciones I y II puede
ocurrir a la vez.
Como resultado de la Fase Lumínica se obtienen
NADPH (gran poder reductor) y ATP (alto contenido
energético) compuestos que serán requeridos en la
siguiente fase y, se desprende O2.
FASE OSCURA
La fijación de carbono se produce en el estroma a
través de una secuencia de 13 reacciones conocida como
ciclo de Calvin. Durante la década de 1950, Melvin Calvin,
Andrew Benson y otros investigadores de la Universidad de
California dilucidaron los detalles de este ciclo. Calvin fue
galardonado con el premio Nobel de Química en
1960.
Las 13 reacciones del ciclo de Calvin se dividen en tres
fases: 1- fijación de CO2 2-
reducción de carbono, y 3- regeneración de
RuBP. Las 13 enzimas que catalizan los pasos del ciclo de Calvin
se localizan en el estroma del cloroplasto. Diez de ellas
también participan en la glucólisis. Estas enzimas
catalizan reacciones reversibles, en las que degradan
moléculas de glúcidos en la respiración
celular y las sintetizan en la fotosíntesis.
1) Fijación de CO2. La primera
fase del ciclo de Calvin consiste en una única
reacción en la que una molécula de CO2 reacciona
con un compuesto fosforilado de cinco carbonos, la ribulosa
bisfosfato (RuBP). Esta reacción está catalizada
por la enzima ribulosa bisfosfato carboxilasa/oxigenasa,
también conocida como rubisco. Es la enzima más
abundante en el cloroplasto, y probablemente una de las
más abundantes en la biosfera. El producto de esta
reacción es un intermediario inestable de seis carbonos,
que de inmediato se descompone en dos moléculas de
fosfoglicerato (PGA), de tres carbonos cada una. El carbono que
originalmente era parte de la molécula de CO2 ahora forma
parte de un esqueleto de carbono; se ha "fijado". El ciclo de
Calvin también se conoce como ruta C3, debido a que el
producto de la reacción inicial de fijación de
carbono es un compuesto de tres carbonos. Las plantas que en un
principio fijan carbono a través de esta ruta se denominan
plantas C3.
2) Reducción de
carbono. La segunda fase del ciclo de Calvin consiste
en
dos pasos en los que la energía y el poder
reductor de ATP y NADPH (ambos producidos en las reacciones
dependientes de la luz) se utilizan para convertir las
moléculas de PGA en gliceraldehído-3-fosfato (G3P).
Por cada seis carbonos que entran en el ciclo en forma de CO2,
seis pueden salir del sistema como dos moléculas de G3P,
que se usarán en la síntesis de azúcares.
Cada una de estas moléculas de G3P de tres carbonos es, en
esencia, la mitad de una molécula de hexosa (azúcar
de seis carbonos). De hecho, se recordará que el G3P es un
intermediario clave en el desdoblamiento del azúcar en la
glucólisis.
La reacción de dos moléculas de G3P es
exergónica y lleva a la formación de glucosa o
fructosa. En algunas plantas, se unen después glucosa o
fructosa para formar sacarosa, el azúcar de mesa
común. La sacarosa puede obtenerse de la caña de
azúcar, la remolacha y la savia del arce. Las
células vegetales también usan glucosa para
sintetizar almidón o celulosa.
3) Regeneración de RuBP.
Obsérvese que, si bien se eliminan dos moléculas de
G3P del ciclo, permanecen diez; esto representa un total de 30
átomos de carbono. A través de una serie de diez
reacciones, que constituyen la tercera fase del ciclo de Calvin,
estos 30 carbonos y sus átomos asociados se reordenan en
seis moléculas de ribulosa fosfato, cada una de las cuales
es fosforilada por el ATP para producir RuBP, el compuesto de
cinco carbonos con que se inició el ciclo. Estas
moléculas de RuBP comienzan de nuevo el proceso de
fijación del CO2, que culmina en la producción de
G3P.
En resumen, los insumos necesarios para las reacciones
de fijación de carbono son seis moléculas de CO2,
fosfatos transferidos del ATP, y electrones (en forma de
hidrógeno) del NADPH. Al final, los seis carbonos del CO2
pasan a formar parte de una molécula de hexosa. Las
moléculas de G3P restantes se usan en la síntesis
de moléculas de RuBP, con las que pueden combinarse
más moléculas de CO2.
INHIBICIÓN
DE LA FOTOSÍNTESIS
El aparato fotosintético está
regulado, en primer lugar, por la luz, que puede ser un
factor
tanto limitante como estresante
Para entender bien el funcionamiento de la fase luminosa
es preciso conocer cómo regulan las plantas el uso de su
primer fuente, la luz, ya que las plantas, al contrario que
muchas algas, que nadan libres en medios acuáticos, no
pueden moverse para huir de la luz cuando llega en exceso o para
buscarla cuando falta en una zona sombreada.
La energía luminosa que llega del sol y es
absorbida por una planta sólo puede seguir dos caminos o
vías posibles: 1) ser transformada eficazmente en otra
forma de energía acumulable (energía
química) mediante el proceso de conversión
fotoelectroquímica de la fotosíntesis; o 2) ser
disipada por conversión a otras formas de energía
radiante no acumulable (como radiación de luz fluorescente
o radiación de calor).
Estas dos vías están en continuo
equilibrio y en relación inversa, cuando aumenta la
primera (transformación eficaz), disminuye la segunda
(disipación), y viceversa. La tasa global
fotosintética de una planta sana y con suficientes
nutrientes inorgánicos aumenta linealmente desde la
oscuridad con la cantidad de luz (llamada densidad de flujo
fotónico fotosintético, PPFD) hasta llegar a un
nivel de saturación, en el cual la energía sobrante
no puede ser utilizada. El nivel de saturación depende de
las características propias de cada especie o variedad, y
del estado fisiológico y las condiciones nutricionales
específicas de cada planta. El exceso de luz tiene que ser
disipado porque, si no, puede "quemar" la planta. De hecho, en
esto consiste propiamente el fenómeno fisiológico
denominado fotoinhibición, en el cual el daño
provocado por la excesiva luz hace disminuir la tasa
fotosintética de las plantas.
El fotosistema II es el centro
fotorregulador en el que tiene lugar el proceso de
fotoinhibición a nivel molecular.
Como centros de reacción, los fotosistemas pueden
ser comparados con un "reactor" de una central nuclear que
está sometido a una tensión energética muy
grande. El reactor tiene que tener numerosos sistemas de
seguridad que lo apaguen o lo cierren en caso de emergencia, para
evitar que la energía se canalice hacia lugares no
previstos.
Algo así sucede en las plantas con el fotosistema
II, que es un reactor natural fotosintético muy bien
protegido y, también, el que tiene más
probabilidades de ser dañado. De todos los fotosistemas
conocidos, el fotosistema II es el que tiene que realizar un
mayor trabajo energético. Los restantes fotosistemas
conocidos, incluido el fo tosistema I de las plantas, ejercen un
trabajo energético menor. Por ello, se puede decir que el
fotosistema II es el de mayor esfuerzo y el punto débil
del aparato fotosintético, como el filamento de una
bombilla, que es lo que primero se puede romper.
Debido a estas propiedades, el fotosistema II es el
centro fotosensible regulador más importante de la
actividad fotosintética, siendo su fotosensibilidad el
medio de control que tienen las plantas para "encender" o
"apagar" el proceso de transformación de energía
luminosa en energía electroquímica. Cuando la
intensidad de la luz sobrepasa la capacidad de absorción y
transformación de la planta, se desencadena en torno al
fotosistema II toda una serie de mecanismos de desacoplamiento y
desactivación, con objeto de maximizar la
disipación de energía y minimizar el daño
global sobre el aparato fotosintético y, de este modo,
proteger a la planta. Así pues, la especial sensibilidad
del fotosistema II está asociada al disparo de los
mecanismos moleculares de fotoprotección.
A los sistemas de protección se añaden los
sistemas de reparación: para mantener la función a
pesar del daño, la planta cuenta con una maquinaria de
reparación que arregla continuamente el daño
sufrido. De hecho, el fotosistema II resulta dañado con
relativa frecuencia, en sus polipéptido central D1. La
ruptura del polipéptido D1 es una manera de proteger todo
el complejo, a modo de un fusible que evita que todo el sistema
se "queme" y que hace que sólo se necesite un recambio
pequeño. Las plantas minimizan así el daño,
centrándolo en un solo polipéptido, de los
más de veinte que contiene el fotosistema II. La
proteína D1 forma parte del centro de reacción del
fotosistema II y en su estructura molecular se insertan, como
grupos prostéticos, clorofila, feofitina, caroteno y
quinonas, las moléculas implicadas en los primeros pasos
del transporte electrónico. El recambio molecular de D1 se
produce continuamente, incluso cuando las plantas reciben
intensidades de luz normales no fotoinhibitorias. Por ello, esta
proteína tiene una tasa de recambio superior en varios
órdenes de magnitud a las del resto de las
proteínas de la membrana tilacoidal.
Considerando la extensión y el volumen del reino
vegetal, se puede afirmar que D1 es la proteína más
sintetizada en todo el planeta. Tal vez éste sea el precio
que las plantas tienen que pagar por ser capaces de utilizar el
agua como fuente de protones y electrones, genera "residualmente"
una molécula tan peligrosa como el oxígeno
molecular gas (O2).
Cuando los mecanismos de desacoplamiento o de
reparación fallan o no son suficientes para aguantar la
intensidad de energía luminosa que llega a una planta,
ésta sufre el citado proceso fotoinhibición
efectivo, que disminuye su eficacia fotosintética y su
capacidad asimiladora .Estos procesos fotoinhibitorios pueden
disminuir drásticamente la producción vegetal y
suelen estar inducidos por la combinación de un exceso de
luz con distintos tipos de estrés nutricional, como falta
de agua (hídrico), falta de algún nutriente
esencial (nitrato, hierro, etc.), exceso de sales en el suelo
(salino), etc.
La fotorrespiración disminuye la
eficiencia de la fotosíntesis, pero puede proteger a las
plantas del exceso de luz en condiciones de bajo CO 2 en el
interior de la hoja.
Una primera consecuencia de la existencia de la
fotorrespiración es que, además de reducir (junto
con la clásica respiración mitocondrial) la
fotosíntesis bruta a las tasas de la fotosíntesis
neta, se produce un gasto energético importante que
disminuye también la eficiencia del proceso de
asimilación del CO2.
Cabe preguntarse, pues, por qué se mantiene en la
planta un proceso aparentemente sin beneficio y que reduce
notablemente la eficiencia de la fotosíntesis (proceso
básico para la producción y el crecimiento de la
planta) ¿Por qué la selección natural no ha
eliminado este proceso? El descubrimiento de la actividad
oxigenasa de la rubisco (muy posterior al del metabolismo del
glicolato) animó a estudiar a fondo este punto de partida
de todo el gasto fotorrespiratorio, y a conocer las posibles
variantes, los efectos ambientales, etc. Con los conocimientos
actuales se comprende que la oxigenación resulta
"inevitable" para el locus activo de la rubisco, quizás
por la similitud de las propias moléculas de CO2 y O2, y
el ciclo fotorrespiratorio permite recuperar el 75% del carbono
perdido por el cloroplasto en forma de fosfoglicolato. Aunque la
fotorrespiración resulte "inevitable", si es posible
minimizarla aumentando la concentración de CO2 o
reduciendo la presencia de O2 cerca de la rubisco. En efecto,
ciertas especies, frecuentes en zonas cálidas y
áridas, y en general de evolución relativamente
reciente, han desarrollado mecanismos (C4 y CAM) que consiguen
incrementar mucho la concentración del CO 2 en el entorno
de la rubisco, reduciendo así a mínimos la
incidencia de la fotorrespiración.
Por otra parte, en condiciones de elevada
iluminación y bajas concentraciones de CO 2 en el interior
de la hoja (por ejemplo, por un cierre estomático en
respuesta a la se quía o la salinidad), la
fotorrespiración puede ser una vía para utilizar el
exceso de ATP y NADPH generado en los tilacoides y, por tanto,
disipar el exceso de energía, previniéndose
así los daños que se puedan causar en el aparato
fotosintético (fotoinhibición). Su papel
sería, pues, complementario al de los carotenoides, en la
medida en que ayudan a proteger la integridad del aparato
fotosintético e condiciones difíciles de
fotoinhibición.
Inhibición de la fotosíntesis
por acción de los herbicidas
Muchos herbicidas actúan sobre la
reacción de Hill (fase luminosa de la
fotosíntesis). Esto supone que la planta no puede acumular
reservas, aunque la acción de los herbicidas trae consigo
otros problemas que eliminarán a la planta antes de que
tenga tiempo de morirse de hambre. Muchos de estos productos
desvían a los electrones de las cadenas de transporte en
el cloroplasto y se acaban formando radicales de oxígeno
muy reactivos que destruyen la clorofila, así como las
membranas celulares. Además, provocan la
destrucción de los carotenoides, y la energía no
recibida por éstos se transfiere a moléculas de
oxígeno, a partir de las que se formarán radicales
sencillos de oxígeno que pueden destruir más
membranas. En resumen: aunque estos herbicidas actúen
sobre la fotosíntesis, su efecto más rápido
y letal suele ser una alteración de la permeabilidad
celular por destrucción de membranas.
Conclusión
Todo el proceso de la fotosíntesis en realidad es
un proceso bastante complejo; por lo que en el presente trabajo
nos esforzamos por describirlo de la forma más simple,
para una mejor comprensión del lector, y resaltar aquello
que interesa a un alumno de Ingeniería
Agronómica.
Para entender bien las necesidades, las capacidades, y,
las limitaciones de un vegetal es de vital importancia entender
la fotosíntesis. Por ello concluimos que los conocimientos
que se aportan en esta investigación son fundamentales
para el buen desempeño en esta materia.
La fotosíntesis es el conjunto de reacciones
químicas que posibilitan casi la totalidad de la vida en
la tierra, ya que es la única ruta para fijar el carbono
presente en todas las células vivas.
Bibliografía
– "Introducción a la
Botánica". Murray W. Nabors
– "Fundamentos de Fisiología
Vegetal" – Joaquín Azcón-Bieto, Manuel
Talón. Segunda edición.
– "Biología". Solomon, Berg, Martin.
Octava edición.
– www.ual.es/personal/edana/bot/mh/temas/t8.doc
Autor:
Rubén Antonio Núñez
Ramírez
Catedrática: Ing. Agr. Daniela
Haupenthal
2011 – Itapúa –
Paraguay
INGENIERÍA
AGRONÓMICA
Trabajo de Investigación presentado
a la materia de Fisiología Vegetal
Alumnos regulares del 3° año de
la carrera de Ingeniería Agronómica.
