Tálamo e hipotálamo
Las placas alares forman las paredes laterales y el
suelo del
diencéfalo, en su parte interna se observa un surco
longitudinal que divide a la placa alar en una porción
dorsal o tálamo y una ventral o hipotálamo. La
fusión
de las dos masas derecha e izquierda sobresalen en la luz del
diencéfalo y conjuntamente forman con las conexiones la
masa intermedia o conexiones intertalámicas.
Las áreas nucleares del tálamo forman dos
grupos
nucleares el grupo
talámico dorsal, encargado de la recepción y la
transmisión de impulsos visuales y auditivos, y el grupo
talámico ventral que actúa como estación de
paso.
En la placa alar en su porción inferios los
grupos nucleares se diferencias en diversos núcleos que se
encargarán de controlar funciones de
digestión, sueño, control de la
temperatura y
conducta
emocional. La acumulación de los núcleos en la
porción ventral del hipotálamo forman el
tubérculo mamilar a cada lado de la línea
media.
Eventos celulares en el desarrollo del
Cerebro.
Sobreproducción de Neuronas y
Apoptosis.
La Embriogénesis produce uno a dos veces
más neuronas de las que están presentes en el
cerebro maduro. Durante 24 semanas de gestación, casi
todas estas neuronas han sido producidas. Sin embargo,
subsecuentemente a esto, hay muerte
selectiva de neuronas.
Muerte de la célula
neuronales genéticamente programada (Apoptosis) es un
rasgo de desarrollo celular que está presente en muchas
áreas del cerebro incluido en el Sistema
Límbico. En contraste con la muerte de
la célula que
es el resultado de la lesión o traumatismo, el Apoptosis
requiere síntesis
de proteínas
y, por consiguiente, es un proceso
celular activo.
Existe un factor de crecimiento que interrumpe el
proceso de la Apoptosis, este factor de crecimiento neuronal,
bloquea la muerte neuronal.
Cono de crecimiento Axonal.
Luego de que los Neuroblastos completan su
división comienzan a emigrar a través de una sola
dirección axonal que se conoce con el
nombre de Cono de crecimiento Axonal. La guía del cono de
crecimiento de un Axón, ramales o dendritas es influida
por factores que orientan el crecimiento hacia un foco
determinado.
Sinaptogénesis.
Una vez que el Axón ha llegado a su lugar
determinado sufre cambios morfológicos y
bioquímicos con el fin de convertirse en el área
presináptica. De la misma manera se forma el área
postsináptica con su receptores y mensajeros.
En el proceso de formación de las
Sinápsis, existe competencia para
desarrollar un tipo de sinápsis u otro, esta
competición deja determinados caminos sinápticos
que el adulto presentará de acuerdo a la constitución e influencia
sinaptogenésica.
Plasticidad Neuronal.
Un importante proceso relacionado con el desarrollo y
organización neuronal es la plasticidad. El
cerebro en vías de desarrollo soporta de una manera
diferente las lesiones o traumatismos que el Cerebro de un
Adulto. Los infantes que tuvieron lesión en la etapa
prenatal, postnatal y neonatal muestran una sorprendente
recuperación funcional y pueden terminar con un
mínimo déficit o ninguno.
ANATOMÍA COMPARADA DEL LÓBULO
LÍMBICO
Durante la primera mitad del siglo XX, la investigación del cerebro estuvo encaminada
en la búsqueda de los mecanismos cerebrales relacionados
con la expresión de conductas emocionales. Mucho de los
trabajos relacionados con el Sistema Límbico estaban
vinculados a la caracterización electrofisiológica
de las áreas neuroanatómicas implicadas en la
expresión de respuestas conductuales que ocurren durante
la defensa de un organismo ante un peligro inminente, respuestas
conductuales durante el apareamiento sexual, conductas de
agresividad, y la expresión de conductas relacionadas con
la ingesta de alimentos,
maternidad, etc.
Todas estas respuestas conductuales ocurren como
procesos
fisiológicos que promueven y determinan la sobrevivencia
de las especies en su medio
ambiente. La investigación de la neurobiología
de las emociones
quedó paralizada a mediados de siglo por la emergencia en
la revolución de las ciencias
cognitivas que intentaban vincular funciones psicológicas
con los mecanismos neurales relacionados con procesos cognitivos,
como los mecanismos de percepción
y memoria.
Más aún, otro factor que impidió el
desarrollo de esta ciencia, se
debió al conocimiento
de la neuroanatomía del Sistema Límbico que
implicaba a este sistema como el sustrato neuroanatómico
de las emociones.
El descubrimiento y la conceptualización del
Sistema Límbico, definido como un sistema
anatómico-funcional que permite integrar y procesar la
información de múltiples eventos
emocionales, permitió también, revolucionar el
concepto
mente-comportamiento
donde los neuroanatomistas de principios de
siglo difundieron el concepto de la neocorteza, estructura
cerebral especializada de los mamíferos, ausente en múltiples
vertebrados cordados. Dado que las funciones cerebrales
superiores —Pensamiento,
razonamiento, memoria, abstracción, análisis y la solución de problemas— son procesos especializados en
los mamíferos, particularmente en humanos y algunos
primates que poseen tejido neocortical, deben ser mediados por
una neocorteza altamente evolucionada, pero no por tejidos
corticales primitivos (paliocortex) y áreas cerebrales
aledañas. El Sistema Límbico ha sido previamente
descrito por la interconexión entre el paliocortex o
corteza primitiva y áreas neurales subcorticales (ganglios
basales) que median en cierta forma aspectos primitivos de
comportamiento emocional y mental, esto es las emociones. La
teoría
del Sistema Límbico como la estructura
neuroanatómica relacionada con el procesamiento de las
emociones empezó a decaer a mediados de siglo, cuando se
vislumbró que lesiones inducidas en el hipocampo
(estructura central del Sistema Límbico) en animales de
experimentación y accidentalmente, en seres humanos,
producía deficiencias importantes en el procesamiento de
diferentes funciones cognitivas, como el caso de la
consolidación de la memoria a
largo plazo. Esta situación era incompatible con la idea
original de que la arquitectura
primitiva o filogenética del Sistema Límbico,
incluyendo el hipocampo, estuviese relacionada con el
procesamiento de complejas funciones cognitivas propias de
humanos y primates superiores. Estas observaciones
experimentales, aunadas al hallazgo de que vertebrados no
mamíferos contienen tejido neocortical similar a los de
los mamíferos, conformado por una estructura más
rudimentaria, dieron como resultado la
desfragmentación sobre la distinción de la
estructura-función
que implicaba, la relación de la neocorteza y paliocorteza
con el procesamiento de funciones superiores y las emociones,
respectivamente
Diversos experimentos,
realizados en estas ultimas décadas, han permitido
extender el concepto neuroanatómico del Sistema
Límbico, permitiendo la inclusión de diferentes
núcleos subcorticales del telencéfalo y estructuras
neurales del tallo cerebral. Si bien el Sistema Límbico
puede incluirse como un sistema neurofuncional mediador de las
emociones, la inclusión de diferentes áreas
cerebrales como parte de este complejo sistema queda aún
sin resolverse. Un vasto número de trabajos experimentales
apoyan el papel de diversas áreas del Sistema
Límbico como estructuras relevantes para el procesamiento
de las conductas emocionales, no obstante, es poco entendido
cómo nuestras emociones surgen por la activación de
los circuitos
neuronales que integran el Sistema Límbico.
Estudios recientes de Imagenología funcional en
el cerebro humano, han permitido entender con mayor
precisión cómo trabaja nuestro cerebro en la
génesis y procesamiento de lo que podemos definir como
emociones. Estos estudios hacen hincapié en que
siempre que se emplea la ejecución de una tarea emocional,
diversas áreas del Sistema Límbico son activadas, y
la activación de estas áreas limbicas están
relacionadas por referencia con la mediación de un evento
emocional. Asimismo, cuando una determinada área del
Sistema Límbico es activada en una prueba cognitiva,
específicamente como la tarea de memorizar, se asume que
existe un aspecto emocional en la respuesta cognitiva.
Desde este punto de vista, se puede inferir que la
teoría límbica de las emociones, está
empezando a explicar y redefinir cómo nuestro cerebro es
capaz de emplear complejas funciones cerebrales que utilizamos
los seres humanos en nuestra vida cotidiana, y en la
génesis de las conductas emocionales. Aunque en el
presente, la teoría del Sistema Límbico resulta
arcaico para explicar los circuitos neurales involucrados en el
comportamiento emocional, las ideas originales surgidas desde
principio de siglo pueden ser tan relevantes para explicar en una
forma general la evolución del cerebro y la emoción.
De esta forma, podemos inferir que las emociones están
sujetas a circuitos neuronales filogenéticamente
primitivos que se han conservado a lo largo de la
evolución de las especies mamíferas y que los
procesos cognitivos pueden emplear circuitos neuronales
diferentes y funcionar en forma independiente a los circuitos que
procesan la información y comportamiento emocional en un
organismo. Por lo tanto, existe una nueva revolución en el
concepto neurobiológico de lo que son las emociones y los
procesos cognitivos, y asimismo, quizá estamos en el punto
de abandonar el viejo concepto del Sistema Límbico como el
sustrato neuroanatómico y funcional exclusivamente del
cerebro emocional.
Comunicación en los
chimpancés
En cuanto a la capacidad de lenguaje de
los chimpancés se ha demostrado que estos animales no
disponen de capacidad de aprendizaje vocal
alfabético; Pueden producir sonidos para manifestar
algún tipo de estado
anímico, pero no están sujetos a ninguna clase de
aprendizaje vocal, lo que se refleja en las estructuras
neuronales que controlan las vocalizaciones.
Un psicólogo americano aplicó a los
chimpancés algo parecido al lenguaje de los sordomudos.
Gracias al mismo, fueron capaces de establecer una serie de
signos capaces
de designar diferentes situaciones y objetos. También se
demostró que la capacidad de aprendizaje de símbolos era superior en chimpancés
educados que en chimpancés que no habían sido
educados. Esto revela que el trabajo
educativo facilita y potencia la
capacidad de cognición. Sin embargo, se presenta una duda
ante el estudio del lenguaje en primates: ¿son capaces de
dominar las operaciones de
reconocimiento de una serie de objetos y símbolos de una
manera similar a los procesos que se siguen en la gramática o en la sintaxis? Los estudios
sobre esta cuestión realizados hasta ahora con primates
demuestran que estos animales son bastante deficientes en cuanto
a la
organización sintáctica de la concepción
y percepción del lenguaje y, más aún, en la
producción.
Hay poca evidencia de que la mayoría de los monos
utilice la gramática en sus comunicaciones, aunque estudios recientes
realizados con chimpancés pigmeos demuestran que son
capaces de captar relaciones sintácticas en las frases.
Incluso pueden llegar a comprender determinadas estructuras
léxicas.
Uno de los principales problemas en los estudios
relacionados con las capacidades cognitivas y
lingüísticas de especies no humanas, en este caso los
chimpancés, es que no son capaces de desarrollar una
producción vocal. Esta evidencia condujo a estudiar a los
delfines, que
poseen una capacidad de producción vocal mucho mayor que
los chimpancés. Los delfines son capaces de entender la
gramática de las minifrases formadas con cuatro elementos
básicos de la lengua:
adjetivos, verbos, sustantivos y preposiciones.
Una de las principales cuestiones pendientes de
determinar con los estudios con delfines y chimpancés
consiste en demostrar que sus habilidades sintácticas
pueden aplicarse también en el proceso de
producción y no, como se ha hecho hasta ahora, sólo
en el de percepción. Queda por demostrar que pueden
estructurar de manera eficiente su producción vocal de
acuerdo con modelos
sintácticos. Sin embargo, se ha demostrado que con un
entrenamiento
específico pueden transferir los símbolos
aprendidos de una manera u otra y, después, desarrollan
esta capacidad con símbolos nuevos.
El grito voluntario para generar alarma es un hecho
difícil de demostrar, ya que puede ser que los monos
estén convencidos de que han dicho la verdad, como pasa
con los niños.
La mentira es, pues, una habilidad de adaptación del ser
humano.
Una de las mayores contribuciones de estudios recientes
de Psicología
comparada ha sido demostrar que las habilidades cognitivas no son
exclusivas de la especie humana. También se extienden a
otras especies de animales como, por ejemplo, las palomas.
Estudios recientes realizados con estos animales han demostrado
que son capaces de dominar tareas no verbales de tipo transitivo.
Los mismos estudios han establecido que el cerebro de las
aves presenta
cierta analogía con el cerebro de los mamíferos y,
por tanto, podríamos hablar también de zonas de
Broca y Wernicke en estos animales.
Un ejemplo de operaciones cognitivas y de la capacidad
de conceptuar se realizó con un grupo de objetos
esféricos y no esféricos en palomas entrenadas para
distinguir un determinado tipo de objetos y palomas entrenadas
para distinguir otro. Cuando aprendieron a identificar los ya
conocidos se les enseñaron nuevos objetos. El porcentaje
de aciertos fue muy elevado, lo que demuestra la gran habilidad
de las palomas a la hora de reconocer objetos desconocidos.
Durante el estudio, las palomas fracasaron cuando los objetos
eran totalmente transparentes.
Sistema Límbico en el cerebro de las
Vacas.
¿Qué hace que una vaca tenga miedo de la
gente? Los animales tienen excelente memoria de sus experiencias,
tanto buenas como malas. Investigaciones
de Joseph LeDoux, de la Universidad de
Nueva York, sobre el Cerebro de animal, han demostrado que los
animales tienen recuerdos imborrables sobre experiencias de
miedo. La memoria del miedo se ubica en un lugar del cerebro
llamado las amígdalas, perteneciente al Lóbulo
Límbico), que son la parte inferior y más primitiva
del cerebro, debajo de la corteza.
Los recuerdos ligados al miedo son permanentes. En los
tiempos en que los bovinos eran animales salvajes, era más
probable que sus predadores se los comieran si no recordaban
adonde habían visto a un león. Con el tiempo, los
animales pueden aprender a superar un recuerdo del miedo, y
asustarse menos del lugar en que tuvieron una experiencia
atemorizante. Pero solamente pueden superar ese recuerdo del
miedo, jamás lo podrán borrar. Por ello, el
esfuerzo debe apuntar a prevenir recuerdos del miedo.
Los vacunos, como otros animales, tienden a desarrollar
recuerdos del miedo que están ligados tanto a lugares
malos u objetos prominentes. Es muy probable que adquieran miedo
a un lugar específico, o a una persona
determinada que usa cierto tipo de vestimenta, si ellos
están asociados a una experiencia dolorosa o atemorizante,
su sietma Límbico integrado actúa
preponderantemente.
Es muy perjudicial para la producción de leche que la
vaca adquiriera miedo a la sala de ordeño. Es necesario
que la primera experiencia de una vaquilla en la sala de
ordeño sea buena. La primera experiencia marca una gran
impresión en los animales. Si una vaquilla se cae o recibe
un choque eléctrico la primera vez que entra a la sala, es
posible que desarrolle un recuerdo del miedo asociado a ese lugar
y la secreción de leche se convierta en un evento
terrorífico para el animal vacuno.
El miedo en las Ratas
Investigaciones realizadas con ratas demuestran el
sustancial efecto que tiene la formación de un recuerdo
miedoso. Se colocó a ratas en un laberinto, y se les
permitió explorar todos los pasillos. Si una rata
recibía un choque eléctrico la primera vez que
entraba a un pasillo nuevo, jamás volvería a entrar
al mismo. Por otro lado, cuando una rata entraba a un pasillo
varias veces y siempre hallaba comida, esto le producía un
recuerdo positivo. Si recibía un choque eléctrico
después de la quinta vez de haber entrado, y
todavía encontraba comida, la rata probablemente
seguiría entrando a ese pasillo.
Si un animal tiene una experiencia dolorosa o
atemorizante la primera vez que está en un lugar nuevo, el
recuerdo del miedo estará asociado a ese lugar. Es
evidente la activación del sistema Límbico de la
rata en dicha experiencia dolorosa o gratificante.

Respuestas de miedo en las diferentes
especies
La comprensión del proceso de miedo en el ser
humano es el resultado del estudio del miedo en otros animales.
Las presiones evolutivas han producido un gran número de
modelos de miedo y alarma en las diferentes especies.
Las respuestas de miedo en los animales incluyen
movimientos expresivos, emisión de olores, sonidos, ritmo
cardíaco y frecuencia respiratoria incrementada,
incremento en la defecación y diuresis.
En estado de terror un perro puede agazaparse y vaciar
sus excrementos. En los vertebrados los movimientos expresivos de
miedo o cólera
incluyen: La erección del pelo en los mamíferos,
de las plumas en las aves y las escamas en los peces.
Las especies comparten miedos similares de la misma
manera que comparten los peligros. Las especies terrícolas
temen las alturas y se retiran de los precipicios; también
muestran miedo ante el poco espacio, la poca iluminación, a determinados
alimentos.
El miedo puede ser generado por muchos estímulos
provenientes de animales de otras especies. El roce repentino, la
proximidad, el acercamiento, el movimiento o
el sentirse mirados son potentes estimulantes del miedo en
humanos y otras especies. Numerosos pájaros,
mamíferos y el humano muestran alarma ante configuraciones
en forma de serpientes, independientemente de que hayan tenido
experiencias previas con serpientes.
Conducta Defensiva en los vertebrados.
Los peces y las larvas de anfibios poseen un cerebro
posterior un par de células
gigantes o células de Mauthner, que controlan los reflejos
locales de la médula espinal, responsables de los
movimientos ondulatorios al nadar. Si se golpea en las paredes
del acuario, los peces se dan la vuelta con un movimiento
centellante, se alejan nadando en dirección opuesta al
estímulo. Todo este proceso se encuentra activado por el
octavo nervio craneal que luego contactan con las células
gigantes, hasta llegar a la médula.
Las respuestas defensivas en los pollos cuándo se
encuentran frente a una mofeta disecada y bajo
estimulación eléctrica del Diencéfalo
produce una adopción
de posturas de lucha, amenaza a la mofeta y la embisten. Si la
estimulación persiste, los pollos huyen o emprenden el
vuelo gritando.
Miedo y conductas relacionadas en los
mamíferos.
Diversas teorías
o modelos de ansiedad, integran a un grupo de regiones cerebrales
en la Conscientización de mecanismos que median la
retirada, la inmovilidad, la defensa agresiva, o la
desviación de un ataque.
Las emociones normales o anormales están siendo
influidas marcadamente por el sistema límbico, incluyendo
el hipotálamo, el septum, la amígdala y el
cíngulo; además los núcleos del
tálamo, el locus Ceruleus, los núcleos del
rafé medial y los núcleos dentados e interpositus
del cerebelo. La destrucción o estimulación de
dichas áreas provoca alteraciones ostensibles de la
conducta defensiva.
ANATOMÍA DEL LÓBULO
LÍMBICO
Componentes del lóbulo
Límbico
Componentes corticales.
Giro del Cíngulo.- Localizado sobre el cuerpo
calloso, posee relaciones con el surco del giro del
cíngulo, giro frontal superior, lóbulo paracentral,
giro precuneus, istmo del giro del cíngulo.
El giro del cíngulo recibe aferentes originados
de muchas fuentes. Una
aferencia importante proviene del núcleo talámico
anterior, que se subdivide en los núcleos antero-medial,
antero-dorsal y antero-ventral.
A través del núcleo anterior del
tálamo, o giro del cíngulo puede ser influenciado
indirectamente por otras regiones. El cortex parietal, el
temporal y el pre-frontal emiten fibras para el giro del
cíngulo.



1. inferior horn | 2. hippocampus Ammon's horn) |
3. pes hippocampi | 4. uncus |
5. parahipocampal gyrus | 6. hipocampal sulcus |
7. dentate gyrus | 8. fimbria hippocampi |
9. crus fornicis | 10.calcar avis |
11 colateral eminence | 12. tapetum |
13. subiculum |

1. uncus | 2. dentate gyrus |
3. fórnix | 4. corpus callosum |
5. fasciolar gyrus | 6. hipocampal sulcus |
7. parahipocampal gyrus | 8. colateral sulcus |
El hipocampo es una estructura importante para la
formación de la memoria a largo plazo. Tiene el
tamaño de un dedo pulgar de niño, y se encuentra
localizado en la profundidad de la porción media del
lóbulo temporal.
Por su parte, el hipocampo (que consta de cuatro
regiones funcionalmente diferentes y conocidas por las siglas
CA1, CA2, CA3 y CA4), contiene el haz de fibras musgosas que unen
el giro dentado con la porción CA3 y la vía
colateral de Schaffer, que comunica a ésta con la
región CA1
La información fluye hacia y a lo largo del
hipocampo por medio de tres vías principales:
- la vía perforante que circula desde la corteza
entorrinal a las células granulares del giro
dentado. - La vía de las fibras musgosas que va desde las
células granulares del giro dentado a las piramidales de
la región CA3 del hipocampo. - La vía colateral de Schaffer que proyecta
desde las células de región CA3 a las de la
región CA1.
Está constituida por las siguientes
estructuras:
Asta de Amón.– o hipocampo propiamente
dicho, nombre dado por parecerse a los cuernos del dios carnero,
es una porción del archipalium que se invaginó en
la parte interna de los ventrículos laterales. En su parte
anterior termina en forma de 3 o 4 engrosamientos que se
denominan digitaciones del hipocampo, llamadas pie del
hipocampo.
Giro Dentado.- o Fascia dentada, es una delgada
banda de corteza situada medialmente al Asta de Amón, se
denomina así por la serie de surcos transversales que
muestran su superficie, en su superficie predominan las
células glanulares, las cuales determinan la
formación de dos áreas anatómicas definidas:
la capa granular formada por cuerpos neuronales de neuronas
granulares que ocupa la ¼ parte de la estructura y la capa
molecular formada por proyecciones axónicas de las
neuronas granulares, ocupa las ¾ partes de la estructura
restante.
Giro Supracalloso.- Capa delgada de sustancia
gris que cubre la superficie superior del cuerpo calloso, llamado
también indusium grisium o rudimento del hipocampo; se
continúa lateralmente con la sustancia gris del Gyrus
Cingulate. Contiene 2 cuerdas que se prolongan longitudinalmente
con las estrías longitudinal media y lateral.
El gyrus Supracalloso se prolonga alrededor y cubriendo
el splenium del cuerpo calloso como una lámina
delicada.

1. Frontal forceps
2. Corpus callosum commissural fibers 3. Short
arcuate fibers 4. Occipital forceps 5. Indusium
griseum 6. Medial longitudinal estria 7. Lateral
longitudinal estría
Giro Subcalloso.
Ventralmente a la rodilla del cuerpo calloso, se
encuentra una zona pequeña de la corteza cerebral, en la
parte media de los hemisferios cerebrales.
Llamado también pedúnculo del cuerpo
calloso, es una lámina estrecha, detrás del area
paraolfatoria y debajo del rostro del cuerpo calloso. Se
continúa con el Genu del cuerpo calloso y con el giro
Supracalloso.

Adyacente al Giro Subcalloso y en la vecindad de la
lámina terminal se delimita el área septal que se
continúa con el septum pelucido. A estas zonas confluyen
por un lado el giro del Cíngulo, el giro Supracalloso, las
estrías longitudinales y la estría olfatoria
medial.
Gyrus Cingulate.
Es una estructura muy extensa que es parte importante
del circuito Límbico, varias funciones del giro cingulate
lo relacionan con la regulación de la conducta maternal;
por ejemplo esta región es importante para mantener la
conducta de la alimentación y en la
habilidad de criar jóvenes. También el gyrus
cingulate interviene en actividades mamíferas complejas,
incluso la regulación del dolor y la atención.
Está localizado sobre el cuerpo calloso, entre al
sulcus del cuerpo calloso y el sulcus cingulate.

- Gyrus rectus
- Superior frontal gyrus
- Cingulate gyrus
- Cingulate sulcus
- Paracentral sulcus
- Central sulcus
- Precuneus
- Cuneus
- Lingual gyrus
Corteza Parahipocampal.
Es la corteza que continúa medialmente al giro
dentado, sobre el giro del hipocampo; constituye una
transición entre el archicortex y el neocortex del
lóbulo temporal, citoarquitectónicamente constituye
la corteza 28 de Brodman.
Se localiza entre la cisura del hipocampo y el surco
colateral y se continúa con el hipocampo a lo largo del
borde medial del lóbulo temporal.
Núcleo Amigdalino
Localizado en el lóbulo temporal próximo
al uncus, se denomina así por que se asemeja a una
almendra. Se ubica por delante y por encima de la punta del asta
inferior del ventrículo lateral. Está fusionado con
la punta de la cola del núcleo caudado.
Este complejo nuclear tiene una enorme importancia en el
humano, mono y felino por Mc Lean y Nauta en sus recientes
estudios neuroanatómicos. De hecho, en los primates
proporciona a la amígdala su proyección aferente
única más masiva siendo esta proyección
recíproca. La amígdala dirige sus proyecciones
corticales a la corteza temporal inferior y a la corteza frontal
(específicamente a la corteza orbitaria del lóbulo
frontal). Por tanto, se proyecta en aquellas partes del neocortex
en las que están representados los eslabones finales de la
cascada de datos sensoriales
hacia el sistema límbico. Evidentemente, el complejo
nuclear amigdalino filtra sus aferencias neocorticales: "Es como
si la amígdala participase en las apreciaciones del mundo
que realiza el cerebro".
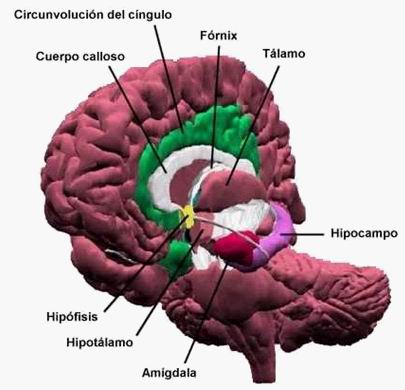
Cuerpos Mamilares.
. Los Cuerpos Mamilares o Tubérculos Mamilares
como Cajal los denominó, forman parte del
hipotálamo posterior y es una de las estructuras
diencefálicas del sistema Límbico. Participan en la
regulación de respuestas neurovegetativas y se han
relacionado con la psicosis de
Korsakoff. Entre los síntomas que presentan estos
pacientes se encuentran severos deterioros de la memoria,
recordando mejor aquellos acontecimientos remotos frente a los
hechos recientes, apareciendo también alteraciones
anímicas como la apatía, desgano.
Son dos cuerpos hemisféricos localizados por
detrás del tuber cinereum. Posee un centro de sustancia
gris recubierto por una cápsula de fibras nerviosas
mielínicas.

Corte coronal que pasa por la región ubicada
entre los cuerpos mamilares y el tallo de la
hipófisis
- Hemisferio cerebral
- Cuerpo calloso
- Tracto mamilo-talámico
- Area hipotalámica dorsal
- Núcleo dorso-medial
- Región hipotalámica lateral
- Núcleo supraóptico
- Núcleo ventromedial (VMN)
- Corteza cerebral
- Cisura ínter hemisférica
- Tálamo

Núcleo Talámico Anterior
Este núcleo está conectado con la
formación reticular, la sustancia nigra el cuerpo
estriado, la corteza premotora y con los otros núcleos
talámicos. Dado que este núcleo está entre
las áreas motoras de la corteza frontal y el cuerpo
estriado posiblemente influirá en la
motricidad.

HISTOLOGÍA
Santiago Ramón y
Cajal, hoy está considerado como el neurocientífico
que contribuyó con mucha dedicación el desarrollo
de las Neurociencias. Su teoría neuronal, demostrando la
independencia
morfológica y funcional de las células nerviosas,
es la base sobre la que se han construido los conocimientos que
actualmente se tienen sobre el funcionamiento de los centros
nerviosos.
Entre 1 887 y 1889, Cajal trabajó " ya no con
ahínco sino con furia", en la elaboración de sus
teorías neurológicas, al igual que otros
histólogos de gran renombre de la época, Cajal
aportó lo más valioso: una genial interpretación funcional de las estructuras
que observó con su microscopio.
Siempre mostró un gran interés
por correlacionar la estructura y la función, aquello le
condujo a decisivas interpretaciones sobre las neurociencias, las
cuales al paso del tiempo le ha dado toda la
razón.
La existencia de puntos de contacto entre los axones,
dendritas y cuerpos celulares, que Cajal describió en su
Teoría neuronal, fueron confirmados mediante el
microscopio electrónico sesenta años más
tarde. Los registros
electrofisiológicos con los que hoy cuentan los
laboratorios de fisiología, también han confirmado
la validez de su Ley de la
polarización dinámica del impulso nervioso, primera
aportación que se hizo para explicar algo que hasta para
el profesor Cajal
le resultaba enigmático: el camino que sigue el impulso o
corriente nerviosa en su tránsito por el sistema
nervioso.
En la época de Cajal no existían
instrumentos que permitiesen la comprobación
fisiológica de esta Ley, pero para el maestro, estaba muy
claro que la corriente nerviosa tenía que seguir una
dirección determinada; el viaje de las dendritas al cuerpo
neurona! y de
éste al axón, que a su vez, transmite el impulso a
otras dendritas de otras células. Fue una genial deducción basada en su Teoría de la
independencia de las células nerviosas. Hoy conocemos que
los puntos de contacto que describió Cajal corresponden a
las sinapsis, lugares en que intervienen diversas sustancias
químicas, que luego los llamaríamos
neurotransmisores.
Cajal desarrolló la Teoría del
neurotropismo, para explicar cómo los axones de las
neuronas en desarrollo
embrionario emigran hacia una dirección determinada
atraídos por sustancias neurotrópicas y que hoy
conocemos como factores del crecimiento. A estas aportaciones
básicas sobre la estructura y función de las
células nerviosas, hay que añadir el estudio
sistemático y detallado, de todo el sistema nervioso
central y periférico.
En 1892, Cajal publicó en la revista
alemana Archiv für Anatomnie und Phisiologie y en la
francesa Bulletin Médicale una serie de conferencias que
había disertado en la Academia de ciencias médicas
de Cataluña con el título "El nuevo concepto de la
histología del sistema nervioso". Animado
por el éxito
de estas publicaciones, dio a la imprenta un
trabajo
más extenso titulado: "Les Nouvelles ideés sur la
structure du systéme nerveux chez l’homme et chez
les vertebrés" (1894), monografía que produjo mucho interés
y difusión en la comunidad
científica. Posteriormente Cajal recopiló sus
publicaciones de quince años de fructífero trabajo,
y así fue como nació el libro "Textura
del sistema nervioso del hombre y los
vertebrados" (Madrid
1899-1904) que es el tratado más completo que se ha
escrito sobre la estructura microscópica del tejido
nervioso.
Todos los vertebrados poseen un sistema nervioso
central, en el cual se distinguen agrupaciones de cuerpos
neuronales (denominados genéricamente núcleos o
centros nerviosos), separados por haces de fibras (denominados
tractos), que conectan a unos centros con otros o con estructuras
periféricas.
Una de las características propias del sistema
nervioso es la de ser un tejido especializado en la
comunicación rápida entre zonas distantes de un
organismo. Es un auténtico sistema de comunicaciones con
capacidad de controlar la captación de estímulos,
procesar la información y elaborar respuestas adecuadas a
estos estímulos. Además, y quizás la
propiedad
más importante, es capaz de aprender, es decir, tras
procesar la información que recibe, es capaz de ajustarse
y modificarse para futuros patrones de actividad.
Entre los numerosos enfoques con los que se estudia el
sistema nervioso (como son por ejemplo la neurofisiología,
la neuroquímica, o la neuropatología), la
neuroanatomía ha resultado ser uno de los pilares
básicos para comenzar a entender cómo funciona
dicho sistema. La neuroanatomía no sólo estudia la
estructura de los diferentes centros nerviosos, sino
también el tipo de conexiones que se establecen entre unos
centros y otros, es decir cómo se comunican unas regiones
del sistema nervioso con otras.
Para establecer los circuitos neuronales es necesario
conocer el origen, la naturaleza
química y
el tipo y disposición de los contactos sinápticos
que llegan a una determinada neurona o grupo de neuronas.
Igualmente, es importante conocer la morfología, la naturaleza química y
las salidas sinápticas de las neuronas que reciben las
conexiones desde el centro de origen. Así, para conocer
los circuitos de un área particular del cerebro (por
ejemplo la neocorteza o el hipocampo) es necesario combinar
diferentes aproximaciones metodológicas en un mismo animal
de experimentación.
Las primeras descripciones detalladas de las conexiones
cerebrales datan de principios de siglo (mucho antes de que se
conociera la actividad de las neuronas individuales), y se deben
a los trabajos del considerado padre de las modernas ciencias del
cerebro, Santiago Ramón y Cajal (Histología del
Sistema Nervioso del Hombre y los Vertebrados, 1911), que con
técnicas de impregnación
argéntica describió las principales vías de
comunicación entre las distintas regiones
cerebrales. Aportó numerosas pruebas para
demostrar que las interconexiones increíblemente complejas
entre las neuronas no se hacían al azar, sino que eran muy
específicas y estaban altamente estructuradas. Cajal
identificó y clasificó en cada caso las diferentes
neuronas, demostrando en ocasiones, hasta donde sus métodos le
permitían, de qué modo se hallaban interconectadas
las mismas.
Ya en los años cincuenta, uno de los
métodos que se emplearon más a fondo para
cartografiar las conexiones entre las distintas estructuras
nerviosas fue el método de
la degeneración. Esta técnica se basa en que los
axones degenerados poseen diferente afinidad tintorial que los
axones intactos. Así, si se destruye un grupo de neuronas
por medios
mecánicos, eléctricos, o por el calor, el
tracto nervioso que procede de ellas degenera (el proceso se
denomina cromatolisis), y antes de desaparecer por completo,
puede ser teñido de manera diferente respecto a sus
vecinos normales. La presencia de axones teñidos
selectivamente en otra región significa que esta segunda
región recibe fibras nerviosas de la parte destruida. Este
método ha posibilitado la obtención de una mapa
relativamente detallado de las principales vías de
conexión en el cerebro, pero la escasa de sensibilidad del
método y los problemas de interpretación de los
datos son los principales obstáculos a esta
técnica.
En los últimos veinte años se han desarrollado una
serie de nuevas técnicas encaminadas a descifrar con
más detalle los circuitos neuronales y que han aportado
avances significativos. Entre estas técnicas se pueden
mencionar:
- Transporte de aminoácidos marcados:
Esta técnica de trazado está basada en las
propiedades fisiológicas de las neuronas. La idea
básica en la que se fundamenta esta técnica es la
de inyectar en una región determinada del cerebro una
sustancia radiactiva (aminoácidos como la leucina o la
prolina marcados con tritio). En dicha región, los
cuerpos celulares toman y la transportan el material
radioactivo a lo largo de los axones. Este transporte
puede ser seguido en el tiempo al registrarse la radiación en una emulsión
fotográfica. De este modo se puede determinar con cierta
precisión la vía que siguen los axones
procedentes de las neuronas situadas en los lugares de
inyección.
2. Trazado con marcadores retrógrados y antero
grados: La idea se fundamenta en la anterior propuesta, pero
tienen la ventaja de no utilizar sustancias
radiactivas.
2.1 Marcaje con sustancias transportadas
retrógradamente: El neuropatólogo sueco Krister
Kristenson descubrió que la enzima peroxidasa de
rábano (en inglés
HRP) era una sustancia transportada retrógrada que se
dirigía por los axones. La técnica comienza con la
inyección de HRP en una determinada región del
sistema nervioso. La HRP es tomada principalmente por los
terminales axónicos y es conducida retrógradamente
hacia los somas. Transcurrido un determinado tiempo (que se
determina experimentalmente dependiendo del animal y de las
condiciones de inyección), el tejido se procesa
aprovechando la actividad enzimática de la peroxidasa: si
se le añade un substrato (peróxido de hidrógeno) a la enzima, junto con un
cromógeno soluble (diaminobenzidina o
tetrametilbenzidina), se da una reacción de
óxido-reducción donde el peróxido de
hidrógeno es reducido, y el oxígeno
liberado se combina con el cromógeno resultando un
pigmento visible tanto a microscopía óptica
como electrónica. Con esta reacción
histoquímica, los cuerpos neuronales marcados con HRP
revelan hacia dónde mandan sus axones. Actualmente existe
toda una familia de
trazadores retrógrados basados en la HRP.
2.2 Marcaje con sustancias transportadas
anterógradamente: Se suele utilizar una lecitina,
obtenida de la judía, denominada Phaseolus
vulgaris-leucoaglutinina o PHA-L o la biocitina (una lisina
biotinizada). Estas sustancias son inyectadas y son tomadas
principalmente por el cuerpo celular y las dendritas. Tras un
tiempo de supervivencia adecuado, se detecta la presencia de
PHA-L mediante anticuerpos que la reconocen. Este procedimiento
revela detalles morfológicos, estudiables a
microscopía óptica y electrónica, de las
proyecciones axonales que no podían ser visualizados con
otros métodos de marcaje.
3. Mapeo de circuitos con virus herpes a:
La técnica se caracteriza por la capacidad de estos virus
para pasar transneuronalmente y replicarse en poblaciones de
neuronas conectadas sinápticamente. El virus es
transportado a través de vías
multisinápticas y la concentración intracelular de
virus aumenta progresivamente con el tiempo. Su distribución por el tejido nervioso se pone
de manifiesto utilizando anticuerpos que reconocen a los virus.
Aunque las condiciones experimentales de esta técnica
requieren muchas precauciones, una de las ventajas de utilizar
virus herpes para identificar circuitos neuronales se basa en la
asunción de que estos virus se replican dentro de las
uniones sinápticas entre neuronas, y que la
infección es consecuencia del paso transináptico
del virus más que de la liberación lítica
del virus al espacio extracelular.
Cada una de estas técnicas han permitido elaborar mapas
topográficos ordenados de interconexiones neuronales
cerebrales; ahora bien se pueden utilizar una "mezcla" de
técnicas.
El
conocimiento de los circuitos neuronales mediante las
técnicas descritas anteriormente han permitido, hacer
estudios evolutivos comparando determinadas vías de
conexión entre los distintos vertebrados amniotas
(reptiles, aves y mamíferos), y que han llevado a la
creación de nuevas hipótesis sobre cómo eran las
conexiones en los vertebrados ancestrales a dichos amniotas
actuales.
Estructura histológica del
Hipocampo.

El archicortex del Asta de Amón está
constituido por 3 capas de células , una más
superficial de células polimorfas, una intermedia de
células piramidales y una profunda o molecular, de
células pequeñas más próxima a los
ventrículos.
Entre las células son características las
células piramidales que presentan un doble penacho de
dendritas, en su base y porción apical.
El álveus.
Es una capa fibrosa que recubre la corteza del Asta de
Amón; hacia la parte media confluyen para formar la
fimbria.
La Facia Dentada.
Tiene también tres capas estructurales, una capa
de células granulares cuyos axones establecen
sinápsis con las dendritas apicales de las células
piramidales del hipocampo, por aquello se la considera una
corteza de asociación.
Núcleo Amigdaloide.
Es en verdad un complejo de varias masas nucleares, que
se agrupan en una porción del uncus del lóbulo
temporal y que se dividen en dos grupos el corticomedial y el
basolateral.
Las células del grupo corticomedial reciben
fibras del bulbo olfatorio; el grupo basolateral está
directamente relacionado con la corteza periamigdalina.
CONEXIONES DEL
HIPOCAMPO
Conexiones Aferentes del hipocampo.
1.-Fibras que se originan en la circunvolución
del Cíngulo y pasan hacia el hipocampo.
2.-Fibras que se originan en los núcleos
septales( núcleos que se ubican dentro de la linea media
cerca de la comisura anterior ) y pasan por detrás del
Fórnix hacia el hipocampo.
3.-Fibras que se originan en un hipocampo y atraviesan
la línea media hacia el hipocampo opuesto en la comisura
del Fórnix.
4.-Fibras provenientes del Indusium griseum que pasan
posteriormente en la estría longitudinal hacia el
hipocampo.
5.-Fibras que provienen del área entorrinal o de
la corteza asociada olfatoria y pasan hacia el
hipocampo.
6.-Fibras que se originan en las circunvoluciones
dentada y del parahipocampo y se dirigen hacia el
hipocampo.
Conexiones eferentes del Hipocampo.
1.-Fibras que se dirigen hacia atrás hasta la
comisura anterior para entrar en el cuerpo mamilar, donde termina
en el núcleo medial.
2.-Algunas fibras que se dirigen hacia atrás
hasta la comisura anterior para terminar en los núcleos
anteriores del tálamo.
3.-Algunas fibras se dirigen hacia atrás hasta la
comisura anterior para entrar en el tegmento del
mesencéfalo.
4.-Fibras que se dirigen hacia delante hasta la comisura
anterior para terminar en los núcleos septales, el
área preóptica lateral y la parte anterior del
hipotálamo.
5.-Fibras que se unen con la estría medular del
tálamo para alcanzar los núcleos
habenulares.
Principales circuitos asociados al sistema
Límbico
Hipocampo-trígono-núcleo mamilar- tracto
mamilotalamico- núcleo anterior del tálamo
proyección hacia corteza cingulada –
proyección hacia corteza Parahipocampal- proyección
hacia el hipocampo.
Amígdala y sus conexiones.
En la amígdala se describen tres grupos nucleares
a) el grupo medial que recibe aferencias del tracto olfatorio y
estría terminal, para luego proyectar hacia área
septal e hipotálamo b) el grupo basal lateral que recibe
aferencias de la corteza temporal y proyecta hacia el
hipotálamo y tálamo ( núcleo dorso-mediano)
y c) grupo nuclear central que recibe aferencias del tronco
encefálico ( núcleo solitario y parabraquial ) y
proyecta hacia el hipotálamo y núcleos viscerales
del tronco encéfalo
Vía septo- hipotálamo-
mesencefálica.
El sistema Límbico a través de estos
circuitos permite influir sobre variados aspectos de la conducta
emocional. Por ejemplo puede inducir reacciones de miedo, rabia,
o emociones asociadas con la conducta sexual.
Fórnix.
Los Axones de las células piramidales entran al
alveolus cuya continuación es la fimbria; esto constituye
la primera porción del Fórnix, el cual termina en
diversos núcleos del hipotálamo y en los cuerpos
mamilares. Algunas fibras cruzan al lado opuesto a nivel del
cuerpo del Fórnix, constituyendo la comisura del
hipocampo.
-Fascículo Cerebral Medio.
Sus fibras se originan en la región septal y en
células de varios núcleos olfatorios. De
allí se dirigen caudalmente a través del
área hipotalámica lateral y avanzan al tegmento
mesencefálico. Presenta conexiones con diversos
núcleos hipotalámicos.
-Estría Medular
–Habénula-Fascículo Habénulo
Pendular.
Desde el área septal y la sustancia perforada
anterior parten un grupo de fibras que rodeando al tálamo
dorsomedialmente, van a terminar en los núcleos de la
Habénula. Algunas pasan al lado contrario a través
de la comisura habenular.
Estría Terminal.
Está formada por fibras procedentes del
núcleo amigdalino que luego de recorre el cuerpo caudado
hasta la cola va a terminar al hipotálamo y a la
región septal.
Comisura Anterior.
Este fascículo cruza la línea media por
delante de las columnas del Fórnix, sus fibras se origina
de los núcleos olfatorios, tubérculos olfatorios,
corteza periforme, núcleos amigdaloides y el neopalium del
polo temporal.
FUNCIONAMIENTO.
La organización del cerebro determina finalmente
su función. Existen tres parámetros importantes en
la organización del cerebro: (1) la densidad de
neuronas, (2) el modelo de
ramificación axonal y de las dendritas, y (3) el modelo de
los contactos de la sinapsis. Estas características
empiezan a desarrollar hacia el extremo del periodo de la cresta
de migración
del neuronal al sexto mes de gestación. Aunque la densidad
neuronal y los modelos básicos de crecimiento axonal y
dendrítico persiguen determinados durante los primeros 2 a
3 años el nacimiento, y el remodelando de las conexiones
del sinápticas continúan a lo largo de toda la
vida.
Se podría imaginar a la población de células nerviosas de la
corteza límbica como la población de una ciudad
dividida en tres distritos, con sus tres vías de
comunicación representados por la amígdala, el
tálamo retromedial y el septum.
Existe evidencias que
sugieren que la población de células nerviosas de
la división amigdalar esta involucradas en la
auto-preservación y en el comportamiento involucrado con
la comida y la batalla para conseguir comida. Por otro lado, la
división septal parece estar involucrada en los
sentimientos y estados expresivos que conducen a la sociabilidad
y la procreación de la especie.
L tercera división o corteza cingulata y partes
del tálamo retromedial se definirá como la
división talamocingular. Esta división abarca
grandes proporciones dentro del cerebro humano, pero no existe la
contraparte en el cerebro reptiliano. Stamm
experimentó sobre la corteza cingulata, en 1955, y
encontró que en ratas hembras adultas, la
destrucción de la corteza cingulata, pero no de la
neocorteza adyacente, producía un déficit en el
comportamiento materno; Cuando se intervinieron
quirúrgicamente a hámster en el primer o segundo
día postnatal, eliminando la neocorteza, pero manteniendo
el Sistema Límbico, estos animales se apareaban
normalmente y cuidaban de sus hijos; Si, se destruía una
gran parte de la circunvolución cingulata junto con la
neocorteza, había déficits en el comportamiento
materno.
El área cingulata anterior y neocorteza frontal
medial adyacente, es necesaria para producir
espontáneamente el llamado de separación. El
llamado de separación puede representar la
vocalización más primitiva y básica que
sirve para mantener el contacto entre madre e hijo.
Parecería que el Gyrus cingulata, que comprende
la parte evolutivamente más nueva del Sistema
Límbico, presenta una combinación de los tres tipos
de comportamiento que caracterizan la transición evolutiva
desde reptiles a mamíferos. Estos son : la crianza en
conjunto con el cuidado materno, la comunicación
audiovocal para el mantenimiento
del contacto madre-hijo y el juego.
La amígdala controla la agresión y las
emociones; Además, la amígdala responde a una gran
variedad de estímulos sensoriales, como sonidos, visiones
y olores; como también estímulos relacionados al
miedo y las emociones.
El hipocampo es el principal responsable del aprendizaje
y la memoria; un daño en
el hipocampo determina una inhabilidad para almacenar la
información nueva en la memoria.
Las funciones del lóbulo límbico
están relacionadas con las bases neuroanatómicas de
la emoción, siendo de orden más jerárquico
en los paleo mamíferos (Delgado-García,1992) y
corresponde al telencéfalo de los reptiles (Curtís
y Barnes,1993). Sin negar el carácter emocional de este lóbulo
con conexiones olfatorias, incorpora una base primitiva de
nuestra afectividad englobando buena parte de las sensaciones
viscerales afines a la misma, por lo que también se ha
hablado de un cerebro visceral (Sanides,1968). Este concepto fue
ampliado por James Wenceslas Papez en 1937 (Donovan,1977;
Delgado-García,1992). Sin embargo, la noción
contemporánea que se tiene sobre el lóbulo
límbico es muy diferente, ya que el continuo estudio
experimental y clínico han desbordado ampliamente los
límites
morfológicos y funcionales, tracendidos en las conexiones
neuronales y en los aspectos bioquímicos en cuanto al
papel de los neurotransmisores en los diversos circuitos
neuronales (Cruz,1987).
En 1937 hubo una gran avance en relación con el
Sistema Límbico, cuando J.W. Papez, planteó la
hipótesis de que
las estructuras del lóbulo Límbico serían el
sustrato neuronal fundamental de las emociones. Más tarde,
Papez fue influenciado por experimentos que sugerían que
el hipotálamo desempeña un papel crítico en
las emociones, y que además tenían un componente
cognitivo por lo tanto una experiencia subjetiva, necesita de la
participación de la corteza y de los circuitos del
hipotálamo.
El circuito de Papez respondió a la
interrogación de cómo los centros corticales se
comunican con el hipotálamo. De acuerdo con este esquema,
las influencias corticales son enviadas al hipotálamo a
través de proyecciones del Giro del Cíngulo a la
formación hipocampal. El hipocampo procesa la
información que llega y la proyecta al Fórnix y a
los cuerpos mamilares del hipotálamo. El hipotálamo
a la vez recibe la información del tálamo a
través del fascículo mamilotalamico y de ahí
al giro del cíngulo. De este va para el giro
Parahipocampal y el hipocampo cerrando el circuito.
Los impulsos nerviosos que circulan por el sistema
Límbico, pueden actuar sobre otras partes del neuroeje,
para ello existen vías eferentes que en conjunto
constituyen las conexiones extrínsecas o de salida del
circuito Límbico. Teniendo en cuenta su punto de destino,
se pueden clasificar de la siguiente forma
Salida neocortical: Toda la neocorteza recibe
fibras procedentes del circuito Límbico, pero
preponderantemente existe una zona preferencial muy importante
que es el lóbulo prefrontal. Estas conexiones
Límbico-corticales permiten que las actividades del
sistema influencien sobre las actividades desarrolladas por la
neocorteza así nos permiten comprender, por ejemplo, como
las emociones pueden repercutir sobre el "pensamiento".
También podemos entender que las emociones, por intermedio
de las vías motoras, que nacen en las áreas motoras
neocorticales, modifique en estado de músculos voluntarios
y así expresarse a través del gesto, la
mímica, la actitud,
posicionamiento.
Salidas medular y tronco-encefálica: a
través de estas conexiones se transportan los impulsos
hacia los núcleos vegetativo (simpáticos y
parasimpáticos) localizados en el tronco del
encéfalo y la medula espinal, como es sabido de estos
núcleos arrancan axones que inervan los vasos y las
vísceras.
Salida Hipofisaria: Que es una de las piezas
claves del sistema Límbico, puesto que existen una serie
de núcleos, cuyos axones terminan en las paredes de los
capilares del lóbulo posterior de la hipófisis por
medio de esta vía la actividad del circuito Límbico
modifica la "formula endocrina" y puede así ejercer
influencia sobre múltiples funciones
orgánicas.
Sistemas de gratificación y de castigo del
circuito Límbico
En 1954 James Old y su alumno Peter Minner, estaban
realizando estudios en ratas sobre la relación existente
entre la formación reticular, la
motivación y el aprendizaje,
para ello situaban micro electrodos en la formación
reticular; Tras realizar, las respectivas estimulaciones
eléctricas siempre que el animal entraba en una de las
esquinas de la jaula, se sorprendieron al observar que el animal
volvía rápidamente a la misma esquina
después de una breve salida y así varias veces. Lo
más importante es que estos dos autores, al percatarse de
la importancia de este hallazgo, abandonaron su proyecto primario
y comenzaron a desarrollar lo que se conocería como la
técnica de la auto estimulación eléctrica
del cerebro, implantaron electrodos intracraneales y permitieron
a las ratas que se auto administraran ellas mismas la corriente
por medio de una palanca.
Está técnica consiste en lo
siguiente:
Implantación de electrodos en diferentes regiones
del encéfalo conectados a un circuito eléctrico que
puede ser manipulado por el animal sometido a la experiencia, de
tal modo que puede administrársele "choques"
eléctricos a voluntad. Los resultados obtenidos con esta
metodología se pueden resumir en lo
siguiente:
Luego del paso de la corriente a través de
electrodos implantados en ciertas zonas del encéfalo se
produce en el sujeto una gama de sentimientos agradables de
bienestar, euforia, placer y satisfacción. Debido a ello
el animal de experimentación se administra choques
eléctricos con gran frecuencia ya que obtiene así
una formidable gratificación. A este conjunto de zonas
encefálicas a partir de las cuales se obtienen estos
resultados constituyen el denominado sistema de
gratificación, cuyos componentes principales son: dentro
del sistema Límbico: el tegmento mesencefálico, la
zona lateral de hipotálamo, los núcleos de la
región septal, al núcleo amigdalino, el hipocampo,
la parte anterior de la circunvolución del hipocampo, el
núcleo caudado; además añadimos la corteza
frontal anterior y el polo temporal. Es importante también
el denominado haz medial del prosencéfalo, haz que se
caracteriza por su amplio origen y por la gran cantidad de
lugares donde proyecta.
A diferencia de ello, la implantación de
electrodos, en otras zonas encefálicas y su
excitación provoca sentimientos disfóricos,
desagradables, penosos e incluso dolorosos, a este grupo de
estructuras, también relacionadas con el sistema
Límbico, se la denomina el sistema de castigo o de
aversión, dicho sistema desde el punto de vista
neuroanatómico está peor definido y las partes
más importantes son: ciertos grupos neuronales del
mesencéfalo, la parte medial del hipotálamo, y
neuronas de la circunvolución del hipocampo.
George, M. S. Et al realizaron el siguiente experimento:
en una serie de mujeres de buena salud, se las inyecto siete
veces, separadas por un intervalo de doce minutos, un trazador
radioactivo por vía intravenosa. A cada una de las mujeres
se les solicitó que recordara dos acontecimientos tristes,
dos felices y dos neutros y que intentaran recrear el sentimiento
que les producía. El análisis estadístico de
los datos demostró que los sentimientos de tristeza
pasajeros, van asociados a un mayor actividad en las
estructuras paralimbicas:, incluidas el cortex prefrontal medial
inferior y el cortex cingular anterior en ambos hemisferios,
superficie medial del lóbulo temporal: corteza
Parahipocampal, así como el cortex prefrontal
izquierdo.
Un sentimiento de felicidad pasajero provoca una
activación no significativa del cortex cingular anterior y
de manara mas clara una reducción del cortex asociativo
secundario: este incluye la corteza orbitofrontal, la
región cingulada y el área Parahipocampal , recibe
aferencias sensoriales y envía proyecciones a otras
áreas corticales incluyendo el córtex prefrontal,
así es una vía a través de la cual las
emociones pueden afectar al plan motor.
A través de La historia comprobamos
cómo el estudioso busca en las distintas regiones del
encéfalo el por qué de ciertas manifestaciones, el
cómo de determinados eventos especialmente aquellos que
tienen que ver con el cambio de
carácter y los trastornos de la
personalidad. Siempre se consideró que las lesiones
del neocortex y de sus conexiones con el sistema Límbico
son responsables de dichos trastornos conductuales.
Además de las observaciones anatomoclínicas,
la ciencia ha
buscado un modelo experimental; Ya vimos como Kluver-Bucy
lograron obtener el síndrome ya descrito por ellos. Sin
embargo tanto en los estudios anatomoclínicos como
experimentales vemos la falta de una adecuada distinción
entre lo expresado por el neocortex y por el sistema
Límbico. Dentro de éste último es menester
distinguir el papel del complejo nuclear amigdalino y sus
conexiones extrínsecas e intrínsecas.
Es necesario señalar que, la zarigüeya
overa(1) (Didelphys albiventris), marsupial de nuestro
país, es un excelente modelo para encarar la
investigación morfológica, fisiológica y
fisiopatológica del complejo amigdalino. En tal virtud
estos marsupiales tienen un muy corto período de
gestación(trece días). Este corto lapso hace que
los recién nacidos sean sumamente inmaduros y que terminen
su desarrollo ontogénico dentro de la bolsa marsupial.
Obviamente en esa bolsa no hay intercambio sanguíneo con
la madre por ausencia de placenta. El único contacto
humoral con la madre es a través de la leche.
Entre los procesos ontogénicos que tienen lugar
después del nacimiento se destaca el 90% de la
formación del sistema nervioso. En el desarrollo
encefálico se distinguen tres períodos: uno
embrionario que se extiende desde la concepción al tercer
día postnatal; uno de diferenciación regional que
se extiende desde el cuarto día hasta los sesenta
días; y uno de maduración que se extiende hasta la
separación completa de la madre.
Las características arriba citadas empujaron a
sus investigadores a comenzar un estudio detallado del
encéfalo de embrión de un día (3). Esto nos
abre la posibilidad de seguir día a día el
desarrollo ontogenético del sistema límbico y en
particular el del complejo amigdalino. El cerebro adulto tiene
más o menos la forma de una pera, con grandes bulbos
olfatorios y hemisferios que divergen en la parte posterior
dejando al descubierto los tubérculos
cuadrigéminos. Los hemisferios cerebrales exhiben una bien
definida fisura rinal, la que se extiende desde la punta del
lóbulo frontal hasta la región occipital. Dicha
fisura separa el relativamente reducido neopalio del extenso
paleocórtex
Después de examinar preparados de
orientación sagital y vértico-frontal con tinciones
específicas para mielina y neuronas, nos ha llamado la
atención lo bien delimitada que se encuentra la
formación nuclear amigdalina, así como su similitud
con la formación nuclear amigdalina del hombre . Este
hecho nos sorprendió, pues estamos en presencia de un
mamífero macrosmático, acallósico y
lisencefálinco. Concomitantemente nos llevó a
examinar los núcleos amigdalinos de los mamíferos
desde el armadillo Chaetophractus villosus hasta los primates.
Así comprobamos que la estructura no se mantenía
con iguales características. Llama la atención la
similitud de organización nuclear amigdalina de
zarigüeya y hombre, puesto que la primera se encuentra libre
de influencias de sistemas
elaborativos de alta jerarquía y tan altamente
complejificados como los del hombre.
Observando el corte vértico-frontal de ambos hemisferios,
teñidos con la técnica de Kluver-Barrera se aprecia
la estructura y magnitud de este núcleo, seguidamente se
visualizan las formaciones subnucleares: núcleo (n.)
medial, n. cortical, n. basal, n. basal accesorio, n. Iateral, n.
central y grupos o nidos de pequeñas células que
constituyen las masas intercalares. Además se visualizan
fibras entre las que se destaca la estría
terminal.
Según estudios de Johnston , en un marsupial del
mismo género y
de otra especie, Didelphys virginiana- las principales aferencias
del núcleo amigdalino están dadas por los
fascículos olfatorios y por la corteza piriforme. Su
principal vía de descarga o eferente es la estría
terminal y sus respectivas subdivisiones. Dicha estría lo
conecta al estriado, hipotálamo, hipocampo, área
septal y la formación reticular a través de su
conexión con el fascículo prosencefálico
medial. La formación nuclear amigdalina dc este modelo se
conecta directa e indirectamente con todo el encéfalo o
dicho de otra manera sobre con todo el sistema nervioso de este
animal y por consiguiente sobre variadas manifestaciones
comportamentales.
El sistema Límbico tiene una influencia
importante en la valoración emocional de todas las
percepciones sensoriales y controla nuestra conducta a
través de la motivación. Cuando se producen trastornos
en la función de las estructuras límbicas, esto
podría representar la causa de muchas enfermedades del
cerebro.
Aproximadamente el 20 por ciento de la población enferma
en el transcurso de la vida de una dolencia neurológica o
psíquica que requiere de tratamiento, guarda su dolencia
una relación con la estructura o funcionalismo del
sistema límbico o de áreas del cerebro que se
encuentran o se presume estarían en estrecha
relación funcional a éste. A este grupo pertenecen
las llamadas neurosis,
enfermedades de dependencia (vicios), esquizofrenias,
síndromes depresivos, formas determinadas de epilepsia y
la enfermedad de Alzheimer.
Casi todas las formas de trastornos de la memoria, trastornos
emocionales, síndromes psicóticos con
pérdida de la percepción de la realidad, ideas
delirantes y alucinaciones tienen como fondo trastornos del
sistema Límbico.
-Comportamiento
alimenticio
Las lesiones en los núcleos amigdaloides producen
cambios en las habilidades alimenticias como hiperfagia o ingesta
indiscriminada de toda clase de alimentos; además debemos
recordar que cambios similares pueden observarse en lesiones en
los núcleos mediales del hipotálamo.
Según Palmiter, los investigadores no especifican
certeramente qué región del cerebro era responsable
del deterioro del comportamiento alimenticio y de los otros
déficit observados en ciertos ratones que se negaban a
alimentarse y morían. Palmiter y sus colegas sospechaban
que las neuronas que producen dopamina en el cerebro medio y que
envían sus axones al estriatum desempeñaban un
papel clave en la integración de señales
del cerebro para producir comportamientos específicos.
Pero no sabían qué parte del estriatum
estaría implicada. El estriatum está dividido en
dos segmentos –el caudate putamen y el núcleo
accumbens.
Mark S. Szczypka, en el laboratorio de
Palmiter, intentó restaurar la producción de
dopamina en el caudate, putamen y en el núcleo accumbens,
inyectando virus inofensivos relacionados con el adenovirus que
llevaban los genes para la Tirosina hidroxilasa y la GTP
ciclohidrolasa I. Los virus fueron suministrados por el coautor
Brian A. Donahue, quien se encuentra en Cell Genesys, Inc., en
Foster City, en California.
Asombrosamente, las inyecciones virales restauraron el
comportamiento alimenticio sólo cuando el blanco de las
inyecciones fue el caudate y putamen. El direccionamiento de las
inyecciones hacia el caudate putamen también
restableció la preferencia de los animales por sustancias
palatables, alimentos de alto contenido graso y agua
azucarada. "Especularía que las señales de hambre
se producen porque estos animales están hambrientos la
mayor parte del tiempo", dijo Palmiter. "Esas señales
probablemente estén alcanzando el caudate putamen, donde
interactuarían con las señales sensoriales que
reflejan la vista y el olor del alimento. Sin embargo, nuestra
conjetura es que en ausencia de dopamina, el caudate putamen no
puede conectar esas señales para producir comportamiento
de búsqueda de alimento".
Las acciones de
las Monoaminas sobre el comportamiento alimenticio han sido
esclarecidas en fisiología animal (H. Saltiel 1989, Bruno
JF 1990), sin que se pueda extrapolar a la patología
humana. Se sabe que receptores beta-adrenérgicos y
dopaminérgicos del hipotálamo lateral reducen la
toma de alimentos, sobre todo en detrimento de las
proteínas. Receptores alfa-noradrenérgicos y
serotoninérgicos son antagonistas en el núcleo
paraventricular del hipotálamo medio, reduciendo o
incrementando, respectivamente la saciedad, sobre todo en lo que
se refiere a los hidratos de carbono. En
los últimos años se ha constatado la presencia de
trastornos de la neurotransmisión serotoninérgica
en una gran variedad de cuadros psiquiátricos en los que
se incluyen los trastornos depresivos, la conducta agresiva, la
conducta suicida, el alcoholismo,
las alteraciones de la conducta alimentaría y los
trastornos de angustia entre otros a partir de datos obtenidos
tanto en investigación animal como en investigación
clínica con fármacos que modifican la
neurotransmisión serotoninérgica. Es posible que en
estos trastornos exista una disfunción
neurobiológica compartida. Existen evidencias
neuroanatómicas que demuestran como las neuronas
serotoninérgicas ubicadas en los núcleos del rafe
tienen un efecto inhibitorio sobre la actividad de diversas
áreas cerebrales implicadas en la mediación de la
ansiedad como el locus Ceruleus, la amígdala y el
hipocampo.
-Comportamiento sexual
La actividad sexual del animal es debida a la tasa y
tipo de hormonas
sexuales en la sangre
circulante. En el sistema nervioso existen receptores que al ser
estimulados activan todo un circuito coordinado; así luego
de la
administración de estrógenos existe la
clásica respuesta del estro.
Con la destrucción de la corteza periforme ,
cercana al núcleo amigdaloide, se obtienen animales, gatos
y micos con una sexualidad
exaltada, que intentan la cópula a cada momento, sin hacer
con animales de la misma especie, ni discriminación de sexo.
Dentro del cerebro interno, es decir de las zonas
cerebrales que se suceden al profundizar en el cerebro tras la
corteza, existen diversas formaciones neurológicas
implicadas en la sexualidad, las más relevantes
serían el sistema límbico, el hipotálamo y
el núcleo estriado.
El sistema límbico no es propiamente una estructura del
cerebro interno, sino más bien un conjunto de estructuras
nerviosas que se hallan rodeando el tronco cerebral, como si se
tratara de un anillo, fundamentalmente, está compuesto por
cinco grandes estructuras que se denominan: hipocampo,
amígdala, septum, giro cingulado y campos de
proyección olfatoria.
Se trata de un sistema extraordinariamente complicado,
tanto en lo que se refiere a los vínculos que mantiene
entre sus diversas estructuras internas como respecto de las
conexiones que estas tienen con otras formaciones externas al
sistema límbico, ya que están íntimamente
ligadas a la corteza cerebral, al tálamo y al sistema
reticular.
Las implicaciones de estos sistemas en la sexualidad no se han
aclarado definitivamente, y tan sólo se conocen algunas
relaciones puestas de manifiesto en diversos trabajos
experimentales.
La amígdala parece jugar un papel central en la conducta
sexual, puesto que la estimulación del septo y del
área pre- óptica media daban lugar a una
erección total en animales de experimentación.
Otra formación de gran importancia dentro del sistema
Límbico es el hipocampo, ya que se cree que ejerce una
influencia moduladora de la tumescencia genital, regulando por
tanto la afluencia de volumen
sanguíneo que fluye al aparato genital, lo cual es hasta
la actualidad el índice más fiable para medir la
excitación y la respuesta sexual.
El sistema límbico esta estrechamente vinculado con otra
zona del cerebro interno que, a su vez, parece estar ampliamente
relacionada con la sexualidad, se trata del hipotálamo,
que también está implicado en la esfera de los
instintos, incluso se ha descrito un área del
placer.
El hipotálamo está relacionado con varias
estructuras nerviosas, recibe vías neurológicas de
comunicación, del sistema Límbico y del
tálamo y de él parten vías de
comunicación hasta el tálamo, la médula y la
hipófisis, también está conectado con la
corteza cerebral, y asimismo se relaciona con el sistema nervioso
autónomo, dando lugar a través de éste a la
gran mayoría de las transformaciones fisiológicas
propias de la respuesta sexual normal.
Además el hipotálamo influye sobre las
glándulas endocrinas, y por tanto sobre la
producción y mantenimiento en el torrente circulatorio de
la mayor parte de las hormonas sexuales.
El hipotálamo está implicado en los
centros del placer, llamados así por lo estimulante que
resulta para los animales de experimentación la
estimulación de esta área, a tal punto de que
prefieren dicha estimulación experimental a su comida
favorita, la estimulación de estas áreas
también se ha realizado en seres humanos, con resultados
variables.
Algunos hombres apreciaron tras el experimento una cierta
sensación de bienestar poco definida, en otros casos dio
lugar a estados de autentica euforia.
Para algunos autores los puntos del cerebro superior que
al ser estimulados dan lugar a un aumento del tamaño del
clítoris similar al de la excitación sexual de la
hembra (tumescencia clitoridiana), serían los mismos que
en el macho causan la erección del pene, tal como parece
haberse demostrado en experiencias con animales.
-Temor
Las aprehensiones e inhibiciones naturales en el animal
pueden desaparecer por lesión de la corteza medial del
lóbulo temporal. Experimentalmente se puede comprobar en
micos, que normalmente muestran pánico
frente a las serpientes, como las toman sin temor y aún se
las llevan a la boca luego de una lesión del lóbulo
temporal. Por el contrario en las ratas una lesión de la
región temporal producen un estado de salvajismo que puede
hacerse reversible con la amigdalectomía..
-Ira
Por estímulo de ciertas zonas del
hipotálamo se puede desencadenar una reacción de
ira o rabia en los animales de experimentación. El
neocortex también una acción
represiva sobre esta reacción es decir la
frena.
La destrucción de los núcleos amigdaloides
en micos y gatos producen animales mansos, plácidos y
somnolientos.
-Manifestaciones
vegetativas.
El lóbulo Límbico y sus interconexiones
determinan la vivencia de reacciones neurovegetativas
específicas que son el resultado de la estimulación
del sistema simpático o parasimpático, debido a las
interconexiones de estos con el lóbulo
Límbico.
Las reacciones obtenida por la estimulación de
núcleo amigdaloide, cuya estimulación produce
salivación, masticación, movimientos intestinales,
cambios en el ritmo cardíaco, dilatación de las
pupilas, posturas de expectativa, reacciones de náusea,
huida.
Por estímulo de la región anterior del
cíngulo y de la región orbital se producen
alteraciones cardiovasculares.
NEUROTRANSMISORES DEL LÓBULO
LÍMBICO.
Bases neuroquímicas de las
emociones
Existen varios sistemas neuronales y
neuroquímicos que median los sistemas de
gratificación-castigo, los mas importantes son las
conocidas como vías aminérgicas, sobre todo las
neurona dopaminérgicas juegan un papel critico en el
sistema de gratificación:
Sistema Serotoninérgico.
Las neuronas que contienen serotonina como
neurotransmisor principal se encuentran principalmente en los
núcleos del rafe tronco encefálicos. Las
células de la parte rostral de estos núcleos
proyectan difusamente hacia la corteza, principalmente al cerebro
anterior, las células de la parte caudal proyectan
principalmente hacia la medula espinal.
Sistema
Noradrenérgico.

Los dos núcleos principales
noradrenérgicos son el locus cerúleus (denominado
así por su apariencia ligeramente azulada en los
cadáveres frescos, contiene aproximadamente la mitad del
total de neuronas noradrenérgicas del tronco del
encéfalo y está localizado en la parte rostral de
la sustancia gris póntica ) y las neuronas tegmentales
laterales que están dispersas en la zona
pontobulbar.
La proyecciones noradrenérgicas se pueden dividir
en :
- Ascendentes: hacia el diencéfalo
(tálamo dorsal e hipotálamo), hipocampo, cerebro
y cerebro anterior. - Descendentes: hacia la medula espinal y tronco del
encéfalo, principalmente núcleos
sensitivos.

Sistema
Noradrenérgico.
Sistema Dopaminérgico.
El sistema dopaminérgico lo podemos dividir, en
base a sus proyecciones eferentes en dos grupos:
- Sistema mesoestriatal
- Sistema mesolimbico y mesocortical
Sistema mesoestriatal, se origina en la substancia nigra
y se dirige básicamente a varias zonas estriatales,
básicamente se le relaciona con el control de movimientos
voluntarios, así la destrucción selectiva del mismo
o su degeneración produce Parkinson.
Sistema Mesolimbico y mesocortical; proyecta desde el
tegmento ventral hacia áreas limbicas y corticales, su
función es aun bastante desconocida, aunque se piensa que
participa en diferentes funciones cognitivas y se le considera
lugar de acción de muchas drogas
antipsicoticas y estimulantes ( como por ejemplo las anfetaminas
).

Sistema
Dopaminérgico
 Página anterior Página anterior | Volver al principio del trabajo | Página siguiente  |