- Definición y
composición general de la leche - Los componentes
lácteos - Contenido de minerales en la
leche - Mecanismo de secreción
de los componentes de la leche - Factores que influyen en la
composición de la leche - Nivel de
producción - Estado de la
lactación - Salud de la
ubre - Época del
año - Número de lactancias y edad
del animal - Influencias
nutricionales - Nivel de
alimentación - La calidad de la
ración - Enfermedades
metabólicas - Bibliografía
La leche continua siendo uno de los alimentos básicos de la
nutrición humana, sustentado
por la gran diversidad y asimilación de los compuestos
esenciales que la integran. Según datos de la FAO (2001), cubre mas
del 20% de las necesidades energéticas, 25% de las proteínas y del 50% del
calcio de la población en países
desarrollados.
Es importante destacar que en la misma medida que se
desarrolla la ganadería y se obtiene
rebaños de mayor potencial genético y
especialización productiva, también se incrementan los
riesgos de aparición de
alteraciones en la composición de la leche, asociadas a
ciertas enfermedades, pues se aumentan
las exigencias ambientales, nutricionales y de manejo (Montero,
2001). Referido a las condiciones de explotación de los
rebaños en las condiciones tropicales, estos riesgos pueden
ser aun mayores, al incrementarse los factores de estrés asociados a la
productividad integral y
específicamente a los rendimientos y calidad de la leche.
La combinación de factores como la época de
seca, baja disponibilidad y calidad de los alimentos y animales de mayor
especialización productiva, constituye un factor de riesgo en el equilibrio fisiológico de
los sistemas involucrados en la
síntesis y secreción de la leche (Ponce et al.,
1992).
El problema en cuestión ha suscitado el interés científico en
algunos países y regiones del mundo, en parte por la posible
relación entre estos trastornos y el uso de los indicadores
físico-químicos como fuente potencial de diagnóstico rápido
de los mismos (Alvarez, 1999a y González 2000), y/o como
vía para evitar cambios indeseables en los parámetros
de normalidad establecidos para el pago al productor y el
cumplimiento de la alta exigencia de calidad de la materia prima en la industria para productos como la leche de
larga vida, productos fermentados y quesos madurados.
La mayor parte de los trastornos metabólicos y
fisiológicos conocidos, tales como son el síndrome de
baja grasa, cetosis, hipocalcemia, fiebre de la leche, inicio y
final de la lactación, generan
alteraciones muy especificas en las características
físico-químicas y pueden ser tratados con el manejo y el uso
de sustancias que regulan el ambiente ruminal, el metabolismo en general o
ambos.
Generalmente se reconoce alteraciones en el contenido de
grasa láctea, sólidos totales y menores en
proteína bruta y caseína, pero no en lactosa, minerales, peso específico,
pH, acidez titulable y en las
relaciones entre ellos, excepto cuando se trata de la mastitis, cuyos cambios
también se identifican con alteraciones específicas
(Contreras, 2000). Por ello resulta importante conocer a fondo el
proceso de las síntesis y
secreción láctea y los factores que influyen en su
comportamiento
Palabras claves: leche, bioquímica,
composición, lactación, enfermedades.
2. Desarrollo
Definición y
composición general de la leche.
La función de la leche en la
naturaleza es nutrir y
facilitar protección inmunológica a los mamíferos jóvenes,
pero también ha sido una fuente de alimentación esencial para el hombre desde tiempos
remotos, quien ha desarrollado la especialización productiva
de varias especies (Hoffmeister, 1989).
La composición química de la leche es la que
sigue:
- 87.3% agua (rango de 85.5% –
88.7%) - 3.9 % grasa (rango de 2.4% – 5.5%)
- 8.8% sólidos no grasos (rango de 7.9 –
10.0%): - Proteína 3.25% (3/4 partes de
caseína) - Lactosa 4.6%
- Minerales 0.75% – Ca, P, citrato, Mg, K, Na, Zn,
Cl, Fe, Cu, sulfato, bicarbonato. - Acidos 0.18% – citrato, acetato, lactato,
oxalato - Enzimas – peroxidasa, catalasa, fosfatasa, lipasa
y otras proteínas - Gases – oxígeno,
nitrógeno, dióxido de carbono - Vitaminas – A, C, D, tiamina, riboflavina,
otras.
- Proteína 3.25% (3/4 partes de
La estructura física de dichos componentes, hace que
sea uno de los líquidos más complejos que se conocen,
pues combina propiedades de emulsión, suspensión y
solución acuosa (Hurley, 2000). En el primer caso dada por
la dispersión de los glóbulos de grasa, en segundo
lugar por la suspensión coloidal de las micelas de
caseína, proteínas globulares y partículas
lipoproteicas y por último, debido a la disolución en
agua de la lactosa, proteínas solubles, minerales, vitaminas y otros componentes.
De aquí se deriva la importancia de sus
interacciones.
La glándula mamaria durante la lactación es
uno de los tejidos más activos y diferenciados de los
mamíferos, donde ocurren profundos cambios y procesos metabólicos
involucrados en la síntesis y secreción de los
componentes (Tucker, 1994), pero que dependen en primera
instancia de la digestión y absorción de los alimentos
y del metabolismo en general, de donde provienen la mayor parte
de los metabolitos necesarios para la síntesis de la leche
(Fox, 1992; Amiot, 1994 y Hurley, 2000).
Lactosa
La lactosa es un azúcar estrictamente
específico de la leche, cuya síntesis y secreción
esta íntimamente relacionada con el volumen total de leche que
producen los mamíferos. La glándula mamaria retiene 900
gramos de agua por cada 50 gramos de lactosa sintetizada, siendo
el principal componente osmótico mediante un proceso activo
de extracción de agua hacia las vesículas de Golgi
donde se produce la síntesis (Forsyth, 1989 y Jensen,
1995).
Desde el punto de vista energético, la
síntesis de lactosa consume hasta un 70 porciento de toda la
glucosa circulante en la vaca
lechera, lo que representa una considerable carga metabólica
para los rumiantes (Ponce y Bell, 1984, Mohar, 1992). En su
síntesis participan varios metabolitos como la propia
glucosa o derivados de esta, proceso que esta regulado por un
complejo enzimático conocido como lactosa-sintetaza,
compuesto por dos proteínas: la proteína A, o
galactosil transferasa que se encuentra en diversos tejidos y la
proteína B o alfa lactoalbúmina que es una
proteína sintetizada por las propias células epiteliales del
tejido mamario (Mepham, 1983, Luquet, 1995, Kennelly,
1999).
La síntesis de la lactosa (Figura 1) es posible
solamente en presencia de la proteína A, pero la
reacción exige entonces concentraciones elevadas de glucosa.
La α-lactoalbúmina permite que se realice la
reacción con cantidades de glucosa muy inferiores, por ello
la tasa de síntesis de la lactosa está regulada por
esta última. (Forsyth, 1989 y Jensen, 1995).

Figura 1. Síntesis de la
lactosa
El flujo de alfa lactoalbúmina y su interacción con la
proteína A insertada en la estructura de la pared del
aparato de Golgi promueve la síntesis continua de lactosa.
Se presume que las limitaciones en el precursor glucosa, y en los
aminoácidos necesarios para la síntesis de las
proteínas del complejo lactosa- sintetasa e incluso en
portadores energéticos como el ATP, pudieran condicionar a
este nivel la síntesis de lactosa y por tanto la producción láctea
(Ponce y Bell, 1984).
Compuestos nitrogenados.
Las proteínas de la leche se originan por
síntesis a nivel del retículo endoplasmático
rugoso de la célula epitelial mamaria
y también por el paso de algunas proteínas de la
sangre mediante un proceso de
difusión. Las proteínas específicas son
sintetizadas a partir de la captación de aminoácidos
sanguíneos en un proceso de ensamblaje, similar a otros
tejidos. Dichos aminoácidos provienen esencialmente de la
digestión de la proteína bacteriana y sobrepasante en
el intestino, por lo que en última instancia el papel de
síntesis de proteína bacteriana en el rumen es un
aspecto de mayor importancia en los rumiantes (Clark, 1996). Como
consecuencia de este fenómeno, las proteínas de la
leche son poco sensibles a las variaciones en la calidad de los
alimentos dentro de ciertos rangos (Kenelly, 1999). La cantidad
de aminoácidos extraídos por la mama no justifica
siempre la cantidad de proteínas que son excretadas,
observándose una captación excesiva de aminoácidos
no esenciales (arginina y valina), mientras que otra parte es
sintetizada en la glándula mamaria a partir de otros
aminoácidos (fenómeno de transaminación, por
ejemplo arginina u ornitina), de ácidos grasos o de glucosa.
(Ikonen y Ojala, 1995).
La síntesis que ocurre en el Retículo
Endoplasmático Rugoso (RER), supone que involucra un flujo
de vesículas desde este hacia el Aparato de Golgi donde se
conforma la miscela de caseína. La manera como ocurre el
tránsito del RER al Aparato de Golgi no está bien
definida aún (Kaneko, 1997). Otros estudios consideran que
las cadenas peptídicas pasan a través del lumen, en el
RER directamente hacia el interior del aparato de Golgi o que el
RER se convierte directamente en el Aparato de Golgi. El proceso
de secreción de la micela de caseína ocurre a
través de una pinocitosis reversa donde la proteína es
vertida dentro del lumen del alvéolo. (Kanno, 1990 y Jensen,
1995).
Esta síntesis exige energía para activar las
reacciones, así como una fuente de nitrógeno y depende
sobre todo de la concentración energética de la
ración. Si faltan aminoácidos, la síntesis de
proteínas se detiene y con ella la de lactosa, con lo que la
producción de leche disminuye. Inversamente, si existe mucha
energía disponible la síntesis de las proteínas
estará estimulada, mucho más que la de la lactosa, por
ejemplo, con el ensilado de maíz (Zea mays), se
puede ganar de 1 a 1,5 puntos por ciento de proteína.
(Kenelly, 1999).
La síntesis de las proteínas de la leche
está regulada sobre todo por mecanismos hormonales y
genéticos en especial de las caseínas,
α-lactoalbúmina y β lactoglobulina. (Van Eenennam
y Medrano, 1991). El contenido de nitrógeno en la leche se
distribuye entre las caseínas (76 %), las proteínas del
suero (18%) y el nitrógeno no proteico NNP (6 %) (Jonker
et al., 1998 y 1999) (Ver Tabla 1).
Tabla 1. Concentración de nitrógeno de la
leche

Fuente: Ikonen et al., (1995)
El grupo mayoritario de las
proteínas de la leche son las caseínas, las cuales
existen en la leche como partículas coloidales conocidas
como micelas de caseína. Su función biológica
está en portar grandes cantidades de Ca y P insolubles para
el mamífero lactante en forma líquida, para formar
después un coágulo estomacal logrando así mejor
eficiencia en la
nutrición.
Además del calcio y el fósforo de la miscela,
la misma contiene también citrato y otros iones,
interactuando con su forma soluble. La estabilidad de la miscela
de caseína depende de la presencia de la Kapa-caseína
(CN) en la superficie de la misma, la cual tiene como
función ser una interfase entre las caseínas
hidrofóbicas del interior de la micela y el medio acuoso. En
resumen, cada micela está constituida por un núcleo o
centro hidrofóbico, recubierto por una capa hidrofílica
que no es otra cosa que la estructura de la Kapa-caseína.
(Mao et al., 1992 y Mercier y Vilotte, 1993).
Las proteínas no específicas de la leche
(inmunoglobulinas y albúmina sérica) refieren Sawyer y
Holt (1993), provienen por difusión de la sangre.
Sin embargo, algunas inmunoglobulinas son sintetizadas y
secretadas por la glándula mamaria en los casos de
infecciones como por ejemplo, la brucelosis.
El nitrógeno no proteico presente en la leche es de
origen alimentario o metabólico. Su componente principal es
la urea que representa del 20 al 75 % de esta fracción
nitrogenada de la leche, en concentraciones de 50 a 200 mg de
nitrógeno por litro de leche. La urea de la leche proviene
de la sangre y su concentración es la misma en ambos
líquidos. Es importante señalar que la urea constituye
un importante indicador del balance energía/proteína en
la dieta de la vaca lechera (Hutjen, 1996; Broderick y
Clayton, 1997 y Hof et.al., 1997).
El resto del nitrógeno no proteico incluye
aminoácidos libres entre los que se encuentran
principalmente el ácido glutámico y la glicina,
así como otros compuestos, residuos de la actividad de la
síntesis de la mama, como nucleótidos, bases
nitrogenadas y ácido orótico (Amiot, 1994).
Grasa.
La grasa láctea está compuesta en más del
95 % de su peso en triglicéridos y el resto de ésteres
de colesterol, ácidos grasos libres y fosfolípidos.
(Walstra y Walstra, 1984 y Place y Gibson, 1988). Estos se
sintetizan directamente en la glándula mamaria, a partir de
los precursores básicos (Acetato y β
– Hidroxibutirato).
Los ácidos grasos preexistentes en la sangre son el
resultado de la síntesis que tienen lugar en los diversos
órganos, como por ejemplo el hígado y el tejido
adiposo. El hígado sintetiza sobre todo ácidos grasos
saturados de 16 y 18 átomos de carbono y monoinsaturados de
16 y 18 átomos de carbono (Mohar, 1992).
El organismo animal no sintetiza los ácidos grasos
poliinsaturados linoleico y linolénico (Mohar,
1992).
Para la síntesis de las grasas, la glándula
mamaria utiliza el glicerol y los ácidos grasos activados
(Jensen et al., 1991).
La glándula mamaria sintetiza in situ los
ácidos grasos saturados de hasta 16 átomos de carbono.
Esta síntesis se realiza bajo el control de las enzimas presentes en el
citoplasma celular y tiene lugar por intermedio del malonil CoA,
activado por la coenzima A. En el caso de la síntesis "de
novo", los ácidos grasos volátiles sirven de
precursores y el alargamiento de la cadena se realiza por
adiciones sucesivas de grupos de dos átomos de
carbono, que provienen del ácido acético circulante y
directamente a partir del ácido
β-hidroxibutνrico, hasta que se
combinan 10 átomos de carbono. (Jensen et al.,
1991)
Si el precursor es el ácido acético, se forman
ácidos saturados de número par de átomos de
carbono, que constituyen la mayoría de los ácidos
grasos sintetizados in situ (Dils, 1986; Fox, 1992 y Hurley,
2000).
La glándula mamaria solamente puede sintetizar como
máximo ácidos grasos de hasta 16 átomos de carbono
(ácido palmítico), por lo que este ácido graso se
acumula en la glándula mamaria. Para la elongación de
la cadena desde 16 a 18 átomos de carbono es necesario la
intervención de otro sistema enzimático
(mitocondrial), lo que no se produce en la glándula (Akers,
1990; Jensen, 1995 y Luquet, 1995).
En la mama, la síntesis de los ácidos grasos
se realiza bajo el control parcial de las proteínas, aunque
este control es menos estricto que en el caso de la lactosa (Cant
et al., 2002). Existe pues una conexión de
síntesis que explica las correlaciones positivas encontradas
entre las tasas de materia grasa y de compuestos
nitrogenados. Debe indicarse que la liberación de los
ácidos grasos por hidrólisis de los triglicéridos
de la sangre tiene lugar por la acción de la
lipoproteína lipasa sintetizada por la glándula
mamaria, que se encuentra en los capilares
sanguíneos.
La actividad de esta enzima es muy importante al
comienzo de la lactancia, lo que podría
explicar la elevada tasa de grasa de los primeros días
postparto, así como el que la proporción de ácidos
grasos de cadena larga de la leche sea mayor al comienzo de la
lactancia. (Jensen et al., 1991).
Dentro de la célula secretora mamaria las
pequeñas gotas de grasa son más numerosas en la
porción apical de la misma que en la membrana
basal.
El incremento del tamaño de las gotas de grasa se
debe a la unión de varias de ellas. Cerca de la membrana
apical de la célula grandes fuerzas de atracción
(fuerza Landon- Van der Walls)
conllevan a que las gotas de grasa sean envueltas por la membrana
citoplasmática (Kanno, 1990), hasta que las gotas están
completamente rodeadas por dicha membrana y son liberadas dentro
del lumen alveolar. Durante este proceso, pequeñas
cantidades de citoplasma pueden quedar atrapadas dentro del
glóbulo de grasa. (Wilde y Hurley, 1996).
Vitaminas.
Las vitaminas son sustancias orgánicas esenciales
para todos los procesos bioquímicos del organismo animal. La
leche contiene las vitaminas liposolubles tales como Vitamina A,
D, E y K (Tabla 2). En el caso de los rumiantes, la vitamina A se
deriva del retinol y de los β carotenos obtenidos de los
pastos y forrajes con la alimentación (Amiot, 1994). La
leche es una importante fuentes de vitaminas
hidrosolubles, entre ellas: tiamina (B1), riboflavina
(B2), Piridoxina (B6), Cianocobalamina
(B12), Niacina, Ácido pantoténico y
también pequeñas cantidades de Vitamina C (ácido
ascórbico).
Tabla 2. Contenido de vitaminas en la leche cruda (Fox,
1992).
Vitamina | Contenido por litro de |
A | 400 |
D (IU) | 40 |
E (μg) | 1000 |
K (μg) | 50 |
B1 | 450 |
B2 | 1750 |
Niacina (μg) | 900 |
B6 | 500 |
Acido pantoténico | 3500 |
Biotina (μg) | 35 |
Acido fólico | 55 |
B12 | 4.5 |
C (mg) | 20 |
Contenido de minerales en la
leche.
Los 22 minerales considerados esenciales en la dieta se
encuentran presentes en la leche. Según Kenelly (1999),
estos se pueden agrupar según su presencia en la leche en
tres grupos:
- Sodio (Na), potasio (K) y cloruros (Cl): estos iones
libres están correlacionados positivamente con la lactosa
y junto a esta mantienen el equilibrio osmótico entre la
leche y la sangre. - Calcio (Ca), Magnesio (Mg), Fósforo
inorgánico (P-i) y citrato: Este grupo presenta una
distribución de 2/3
partes del Ca, 1/3 del Mg, 1/2 del P-i y menos de 1/10 del
citrato en forma coloidal presentes en la micela de
caseína. - Sales disueltas de Ca, Mg, citrato y fosfatos: Estos
elementos son dependientes del pH y contribuyen al equilibrio
ácido-básico de la leche.
A pesar que el contenido mineral de la leche es muy
amplio (Tabla 3), desde el punto de vista nutricional, el calcio
y el fósforo tienen la mayor la mayor importancia. (Walstra
y Jennes, 1984; Place y Gibson, 1988 y Hurley, 2000).
Tabla 3. Contenido mineral de la
leche.
Mineral | Contenido por litro de |
Sodio (mg) | 350 – 900 |
Potasio (mg) | 1100 – 1700 |
Cloruro (mg) | 900 – 1100 |
Calcio (mg) | 1100 – 1300 |
Magnesio (mg) | 90 – 140 |
Fósforo (mg) | 900 -1000 |
Hierro (μg) | 300 – 600 |
Zinc (μg) | 2000 – 3000 |
Cobre (μg) | 100 – 600 |
Manganeso (μg) | 20 – 50 |
Yodo (μg) | 260 |
Flúor (μg) | 30 – 220 |
Selenio (μg) | 5 – 67 |
Cobalto (μg) | 0.5 – 1.3 |
Cromo (μg) | 8 – 13 |
Molibdeno (μg) | 18 – 120 |
Nickel (μg) | 0 – 50 |
Vanadio (μg) | Trazas – 310 |
Arsénico | 20 – 60 |
El contenido mineral de la leche es estable y solo se
reportan cambios sustanciales en el caso de los electrolitos,
afectados por el estado de la lactancia y
por enfermedades como la mastitis o cuando ocurren alteraciones
en el tejido mamario (Armenteros, 1998 y Hurley,
2000).
Mecanismo de secreción
de los componentes de la leche.
El mecanismo de secreción láctea puede
resumirse de la siguiente manera (Figura 2)
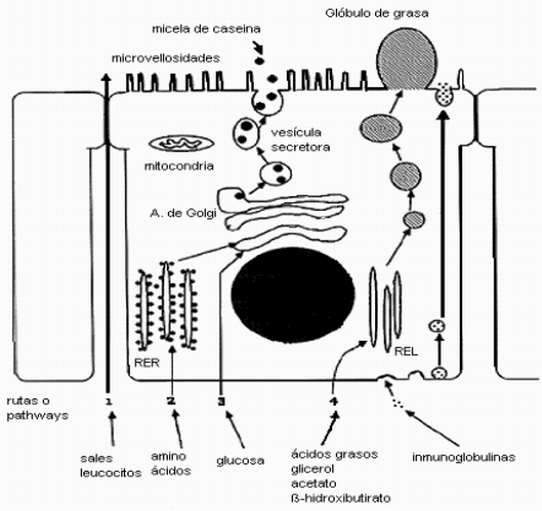
Figura 2. Mecanismo de secreción
láctea
Adaptado de Hurley (2000).
Factores que influyen en la
composición de la leche.
Sobre la composición de la leche influyen factores
nutricionales y factores no nutricionales (Strandberg y Lundberg,
1991 y Durr et al., 2000).
Factores no nutricionales.
Raza.
Existen notables diferencias entre razas con
relación a los componentes mayores de la leche, donde se
distingue la raza Holstein con niveles de sólidos más
bajos si se compara con otras razas como la Jersey, que registra
la mayor composición (Tabla 4).
Tabla 4. Composición media de la leche en las
principales razas lecheras.
Por | ||||
Razas | Grasa (%) | Proteína | Proteína | Sólidos (%) |
Ayrshire | 3.88 | 3.31 | 3.12 | 12.69 |
Brown Swiss | 3.98 | 3.52 | 3.33 | 12.64 |
Guernsey | 4.46 | 3.47 | 3.28 | 13.76 |
Holstein | 3.64 | 3.16 | 2.97 | 12.24 |
Jersey | 4.64 | 3.73 | 3.54 | 14.04 |
Shorthon Lechero | 3.59 | 3.26 | 3.07 | 12.46 |
Adaptado de Amiot, (1994)
La raza constituye hoy uno de los factores más
relevantes a considerar en la composición de la leche,
puesto que la grasa y proteína lácteas son caracteres
genéticos con alta heredabilidad (Imagawa et.al.,
1994).
La heredabilidad estimada para la producción de
leche es relativamente baja (0.25), según Mercier y Vilotte
(1993), sin embargo la heredabilidad estimada para la
composición de la leche es bastante alta (0.50).
Opuestamente los factores ambientales como la nutrición y el
manejo alimentario pueden tener mayor efecto sobre la
producción que sobre la composición de la leche
(Ng-Kwai-Hang et al., 1984 y Ponce, 1984). En la
práctica, en los últimos 20 años se ha logrado un
incremento de los sólidos de la leche, manteniéndose
altos niveles productivos, debido al manejo combinado de la
genética y la
alimentación
Es por ello que la selección genética en
ganado lechero deber ser enfocada directamente al incremento de
los componentes lácteos, aunque por el momento resulta muy
difícil lograrlo puesto que existe una asociación
genética negativa entre estos y la producción de leche
(Blackburn, 1993).
De igual manera, la selección de sementales sobre
la base de porcentajes de grasa y proteína láctea
resultará en una reducción de los rendimientos en
producción de leche y en una mejora mínima en los
rendimientos en grasa y proteína en su descendencia. Debido
a que el pago de la leche está basado en el nivel de
sólidos, se debe enfocar la selección de los sementales
basado en el rendimiento de grasa más proteína
láctea, lo que resultará no solo en un incremento en
producción de leche sino también en mayores porcentajes
de grasa y proteína y en sus rendimientos individuales
(Velmala et al., 1993).
Los rendimientos en grasa, proteína, sólidos
no grasos y sólidos totales son altos y positivamente
correlacionados con la producción de leche. Sin embargo
los valores porcentuales de
los mismos en la composición de la leche disminuyen en la
misma proporción (Beever et al., 1991 y Coulon et
al., 2000).
El concepto del rendimiento de los
componentes lácteos contra la composición de la leche
se puede ilustrar comparando diferentes producciones de leche con
una composición en proteína similar (Mao et al.,
1992). Si la producción o rendimiento por vaca se incrementa
de 29,5 a 31,8 Kg. mientras la composición de la
proteína permanece constante en 3.1 por ciento, se alcanzan
incrementos de 0.07 Kg. más de proteína por día.
Si el porcentaje de la proteína aumenta de 3.1 a 3.2 por
ciento mientras que la producción por vaca se mantiene a 29,
5 Kg, los aumentos de la producción o rendimiento de la
proteína por sólo 0.03 Kg por vaca por
día.
El curso de la lactancia, no solo afecta la
producción de leche, sino también la composición.
Normalmente, un aumento en el rendimiento de leche es seguido por
una disminución en los porcentajes de grasa y proteína
en leche mientras los rendimientos de estos componentes
permanecen igual o en aumento. (Knight y Wilde, 1987; Akers, 1990
y Pérochon et al., 1996).
Los cambios en los rendimientos productivos durante el
ciclo de lactancia, influyen de manera inversa a la
composición. Generalmente, en el primer tercio de la
lactación y concomitante con el pico de lactancia, se
registran las menores concentraciones de grasa, proteína y
sólidos de la leche, situación que se invierte al final
de la lactancia (Akers, 1990, Beever et al., 1991,
Blackburn 1993 y Hurley, 2000). Se exceptúan de este cuadro,
las concentraciones de lactosa y potasio que disminuyen al final
de la lactancia (Ponce y Bell, 1985). Lo descrito anteriormente
puede constarse gráficamente en la figura 3.

Figura 3. Comportamiento de la
producción de leche, el porcentaje de grasa y proteína
durante el ciclo de la lactancia.
Adaptado de Oldaham (1991).
Sin embargo, este comportamiento se ajusta más a
rebaños de vacas altas productoras, donde se aseguran todas
las condiciones de manejo y alimentación, ya que otros
reportes sobre animales de menor producción y en condiciones
rústicas de explotación no se observa este
comportamiento típico (Rearte, 1993 y Gonzáles-Stagnaro
et al., 1998). En estos las curvas de lactancia son menos
pronunciadas y en ocasiones lineales (Ponce, 1984). En tales
condiciones, se evidencia una mayor influencia de los factores de
manejo y alimentación. El tipo de animal y las condiciones
de explotación, también son elementos muy relacionados
con estos factores (Pérochon et al.,
1996).
La mastitis es la enfermedad que más afecta la
producción y la composición de la leche y por ello ha
sido ampliamente estudiada (Calvinho, 1995; Smith, 1996 y
Rajala-Schultz et al., 1999). La tabla 5 muestra los cambios que ocurren
en la composición de la leche con niveles altos de
células somáticas, ocurriendo una reducción en el
contenido de grasa y caseína y un aumento en el contenido de
suero de leche. Estos cambios en las proteínas de leche, en
unión con modificaciones en la lactosa, el contenido del
mineral y pH de leche, tienen como resultado bajos rendimientos
en la producción de queso y alteraciones en las propiedades
y en la aptitud industrial de esa leche (Armenteros, 1998). Bajo
dichas condiciones se aprecia un tiempo de coagulación
más largo y una cuajada más débil que la leche no
afectada.
Tabla 5. Cambios en la composición de la leche
asociados a elevados conteos de células somáticas
(CCS).
Componente | Leche | Leche con |
SNG (%) | 8.9 | 8.8 |
Grasa (%) | 3.5 | 3.2 |
Lactosa (%) | 4.9 | 4.4 |
Proteína (%) | 3.61 | 3.2 |
Proteínas del suero | 0.8 | 1.3 |
Sodio (mg) | 570 | 1050 |
Cloruro (mg) | 911 | 1470 |
Potasio (mg) | 1500 | 1750 |
Calcio (mg) | 1200 | 400 |
Adaptado de Philpot | ||
Los porcentajes de grasa y de proteína son más
altos durante el invierno y más bajos durante el verano
(Dahl et al., 1998). Esta variación está
relacionada con cambios en la disponibilidad y calidad de los
alimentos y las condiciones climáticas. Durante el verano
(época lluviosa) los pastos son bajos en fibra y se deprimen
los niveles de grasa en la leche. Además la alta temperatura y humedad
relativa, disminuyen los niveles de consumo (Dahlet et al.,
1998 y De Lima et al., 2001). Durante el invierno
(época poco lluviosa) disminuye la disponibilidad y la
calidad de los alimentos (pastos y forrajes), por lo que aumentan
los niveles de grasa en leche, pero disminuye la producción
de leche (Coulon y Pérochon, 2000 y White et al.,
2002).
Los factores ambientales en la mayoría de los casos
influyen directamente en el nivel de consumo de los animales
dando como resultados variaciones significativas en la
producción de leche y en la composición (De Lima et
al., 2001). Cuando la temperatura se encuentra por encima de
los 30 °C se reduce la producción de leche, además
de los niveles de grasa y proteína, debido a la
reducción del ingreso de energía a través de la
dieta. La combinación del estrés calórico, con la
pobre suplementación o una dieta basada solamente en
forrajes condiciona en la lactancia temprana y media a una
disminución de los rendimientos lácteos.
Número de lactancias y edad
del animal.
Los niveles de producción de leche aumentan con las
sucesivas lactancias de la vaca, obteniéndose los mayores
volúmenes entre la tercera y la cuarta lactancia, lo que
depende en gran medida de la edad de incorporación del
animal a la reproducción y el manejo
del mismo durante su vida productiva (Ponce 1984, Beever
et.al., 1991, Imagawa et al., 1994, Aranda et
al ., 2001).
Mientras el contenido de grasa en la leche permanece
relativamente constante, el contenido de la proteína en
leche gradualmente disminuye con avance de la edad (Wilde y
Hurley, 1996 y Hurley, 1996). Un estudio del DHIA (2000), en
rebaños Holstein Friesian, indica que el contenido de la
proteína en leche disminuye típicamente de 0.10 a 0.15
unidades sobre un período de cinco o más lactancias o
aproximadamente 0.02 a 0.05 unidades por lactancia.
Número, intervalo y tiempo de
ordeño.
El incremento de uno a dos y hasta tres ordeños
diarios, es un criterio más de tipo práctico y
económico, que de otra naturaleza. Cuando se tienen vacas
con producciones por debajo de los 10 litros de leche, la
realización de dos ordeños diarios no se justifica,
pues los incrementos productivos no rebasan por regla general el
10 – 15 porciento (Ponce 1984, Erdman y Varner, 1995). Una
situación diferente se presenta en rebaños altos
productores y con excelente manejo y alimentación, puesto
que el doble ordeño se hace necesario cuando la presión interna debido al
rápido llenado de la ubre se convierte en un factor
limitante para la producción (Bigner et
al., 1997).
El intervalo entre ordeños, es más común
de 12 horas o con ajuste de 10/14 horas, tratando de evitar las
altas temperaturas durante el verano, potenciando así las
horas de máximo pastoreo (Klei et al.,
1997).
El tiempo de ordeño está relacionado en la
práctica con la rutina empleada, aunque debería
ajustarse más a las características de
ordeñabilidad de los animales. Para ello agrupar vacas de
rápido ordeño (menos de 7 minutos) es una práctica
aconsejable y dejar para el final las vacas que demoran más
durante el ordeño (Lacy-Hulbert et al., 1999). En
todos los casos, la composición láctea está
relacionada inversamente con el número e intervalo en que se
produce el mayor volumen de producción láctea. Sin
embargo un ordeño incompleto aumenta el volumen de leche
residual en la ubre, disminuye el porciento de grasa y la
producción de leche en el siguiente ordeño, lo cual de
suceder de manera continua, logra una depresión irreversible de la
producción láctea (Ponce 1984, Knight y Wilde 1987 y
Lacy-Hulbert et.al., 1999).
Del conjunto de alteraciones en las características
físico-químicas de la leche, la concentración de
grasa es la que resulta más sensible a cambios nutricionales
y puede variar casi 3.0 unidades porcentuales. Los efectos que
tiene la alimentación sobre la concentración de la
proteína láctea puede producir cambios hasta de 0.60
unidades porcentuales (Palmquist, 1993). Las concentraciones de
la lactosa y minerales, no responden previsiblemente a ajustes en
la dieta y tampoco se han reportado efectos sensibles sobre el
pH, la acidez y el peso específico de la leche. (Freeden,
1996 y Mackle et al., 1999).
Las vacas con bajos niveles de alimentación reducen
la producción de leche y el porcentaje de lactosa solo
dentro de ciertos límites, sin embargo se
producen aumentos en el porcentaje de grasa láctea
(Palmquist, 1993). Por regla general, cualquier ración que
aumenta la producción de leche reduce generalmente el
porcentaje de grasa en la leche. La subalimentación es el
único régimen nutricional que altera la lactosa donde
el porcentaje de la misma solo se reduce levemente (Oldaham,
1991).
Diversas investigaciones han demostrado
que por cada 30 Kg de incremento de peso vivo al momento del
parto se logran incrementos en
la producción de leche de 122 Kg, 8 Kg de grasa y 4 Kg de
proteína durante las primeras 20 semanas de la lactancia
(Rearte, 1993). Sin embargo los efectos de la condición
corporal sobre los porcentajes de grasa y proteína son
pequeños.
Durante el primer tercio de la lactación, las
demandas nutricionales de la vaca lechera, son mayores que la
capacidad física de cubrir dichas demandas y ocurre un
proceso de balance energético negativo. (Fredeen,
1996).
Durante la lactación los efectos del nivel de
alimentación sobre el porcentaje de grasa y proteína
son variables (Odahalm, 1991).
Durante la lactancia si se incrementa el consumo de alimentos,
también se incrementa la producción de leche y los
rendimientos en grasa y proteína. Con el incremento del
consumo, el porcentaje de grasa en la leche tiende a disminuir,
sin embargo el porciento de proteína manifiesta un ligero
incremento. Esta última, en animales adecuadamente
alimentados se encuentra raramente por debajo de 3.2 porciento,
sin embargo en rebaños con malas condiciones alimentarias
puede decaer hasta 2.8 porciento (Coulon y Pérochon,
2000).
El uso de pastos de buena calidad en la
alimentación de la vaca lechera trae como resultado un
incremento en la producción de leche y en los rendimientos
en grasa y proteína lácteas (Juárez et al.,
1999 y Pérez, 2001). La influencia de especies de pastos
sobre la producción y composición de la leche se
muestra en la Tabla 6.
Tabla 6. Efecto de algunas especies de pastos sobre
la producción de leche, y la composición de grasa y
proteína.
Experimento I | Experimento 2 | |||
Kikuyu (Pennisetum | Ryegrass (Lolium | Ryegrass (Lolium | Trébol (Trifolium | |
Producción de leche | 13.4 | 19.4 | 16.5 | 18.9 |
Grasa (%) | 3.7 | 3.5 | 3.7 | 3.5 |
Grasa | 0.51 | 0.70 | 0.59 | 0.69 |
Proteína % | 2.9 | 3.2 | 3.0 | 3.2 |
Proteína | 0.40 | 0.64 | 0.51 | 0.62 |
Adaptado de White et al.,
(2002).
Con un aumento en la producción de leche, así
como del porciento de proteína, se produce un incremento
además en los rendimientos en proteína en la leche
(White et al., 2002).
Las diferencias que se presentan entre las especies de
pastos son inherentes al consumo voluntario de los animales.
Cuando los niveles de alimentación son similares entre el
ryegrass (Lolium perenne) y el trébol (Trifolium
repens), dichas diferencias son similares.
En el caso del trópico, García et al.,
(1988), señalan que los pastos constituyen la base
alimentaria de la vaca lechera. En este caso se presenta un bajo
nivel de energía y proteína, lo cual condiciona que
ello sea la principal causa que afecta la producción de
leche y la composición y se hace entonces hace necesario
suplementar con granos o cereales y lograr balancear
adecuadamente la ración.
El empleo de concentrados a base
de cereales, resulta en un incremento en la producción de
leche y en los rendimientos en grasa y proteína (Ponce et
al., 1990). Sin embargo, si se cubren los niveles de
proteínas en la ración total de la vaca lechera, un
suplemento proteico adicional logrará los mismos incrementos
en el porcentaje de proteína láctea que una ración
de cereales con un nivel de energía similar (Kalscheur et
al., 1999).
Cuando el uso de concentrados en la dieta de la vaca
lechera es elevado y constituye más del 60 porciento de la
misma se produce cierta depresión en el porcentaje de la
grasa láctea (Ponce et al., 1990).
Para mantener e incluso incrementar los niveles de
grasa láctea, es aconsejable además el empleo de
pequeñas cantidades de fibra en la dieta. El aporte diario
de 2 a 3 kg./vaca de heno unido a una alimentación a base de
pastos, logran mantener el porcentaje de grasa en la leche
(Balbinotti et al., 2001). Además para eliminar el
efecto de la depresión de la grasa se necesita de un 15
– 17 porciento de fibra en la dieta. (Van Soest et
al., 1991). Sin embargo el mayor contenido de fibra en los
pastos maduros hace que disminuya su calidad (menor
digestibilidad), se afecte en consecuencia el consumo de materia
seca por parte de los animales, y disminuya la producción
diaria de leche (Rearte, 1993).
Cuando las vacas consumen pasturas con mayor cantidad de
fibra, al avanzar la primavera, se recuperan los porcentajes de
grasa en la leche, pero a costa de una menor producción de
leche, obteniéndose menores producciones de grasa diaria. El
menor consumo de energía de las vacas sobre pasturas maduras
se refleja también, en menores porcentajes de proteína
de la leche lo que unido a una menor producción, provoca
importantes disminuciones en la producción diaria de
proteína (Mackle et al., 1999).
El mayor valor nutritivo de los pastos
en la época lluviosa, se debe a que en esta época una
mayor proporción de la proteína de los pastos y
forrajes está como proteína verdadera, mientras que en
la época de seca, parte de dicha proteína está
como nitrógeno no proteico (Oldaham, 1991; Rearte, 1993; De
Lima et al., 2001)
En resumen, los efectos de la alimentación sobre los parámetros físico-químicos son mínimos. Solo pequeños cambios en las concentraciones de lactosa y minerales
han sido reportados (Jensen, 1995), aunque sobre este particular no existen evidencias que reflejen alteraciones en el equilibrio mineral o en la estabilidad térmica de
la leche por estas causas (Ponce, 1999).
Los rebaños lecheros de alta producción deben
contar con un adecuado balance de nutrientes, especialmente en
los períodos de mayores necesidades nutricionales, que se
corresponden con el inicio de la lactación. En el
período inicial, la vaca llega al nivel máximo de
producción y a su vez el consumo voluntario se deprime,
además los aportes de la dieta no logran cubrir los elevados
requerimientos metabólicos, debiendo movilizar sus reservas
corporales para compensar esta situación (Pocknee, 1998 y
Barros, 2001).
La ocurrencia entonces de desbalances nutricionales,
deficiencias o inadecuado manejo de los programas de alimentación
para vacas lecheras pueden conducir a la aparición de varios
trastornos en la salud de los animales los cuales se
denominan enfermedades o trastornos metabólicos de la vaca
lechera (Alvarez, 1999a).
Los trastornos metabólicos cobran mayor importancia
en la misma medida en que la ganadería lechera se
desarrolla, pues una vez establecido un control estricto sobre
las enfermedades infecto-contagiosas, el aumento del potencial
genético de los rebaños y se optimizan las condiciones
de manejo y alimentación se logra aumentar la productividad
lechera (González, 2000). Es entonces cuando comienzan a
aparecer con mayor frecuencia dichos trastornos. Estas
alteraciones constituyen después de la mastitis el principal
problema en todos los países de ganadería lechera
desarrollada, por las grandes pérdidas económicas que
implican (Alvarez, 1999a).
El estrés producido por los trastornos
metabólicos reduce la capacidad de resistencia y comprometen el
funcionamiento del sistema inmunológico del animal
(Contreras, 2000). Si esta situación no se previene a
tiempo, se producen consecuencias altamente costosas en la
reproducción, y la producción de leche (Alvarez,
1999b). En los rebaños lecheros de los Estados Unidos (NRC, 2001), las
pérdidas por concepto de los trastornos metabólicos se
calculan entre un 20 – 25 porciento de las pérdidas
totales anuales.
A continuación se relacionan las principales
enfermedades metabólicas de la vaca lechera.
Síndrome de la vaca gruesa.
Esta patología se presenta durante el período
seco y la lactancia temprana (Barbano, 1999), y reconoce que el
alto nivel energético de la ración produce una alta
acumulación de grandes cantidades de grasa de reserva. Ello
desencadena en la primera etapa de la lactancia una
lipólisis elevada con un aporte excesivo de ácidos
grasos al hígado, conduciendo a una lesión
hepática, con consecuencias metabólicas y
reproductivas, razón por la cual algunos autores la llaman
Síndrome del hígado graso (Kaneko et al.,
1997).
El complejo sintomático descrito está dado por
obesidad, problemas reproductivos tales
como dificultad al parto, retención placentaria, además
trastornos nerviosos como depresión y fatiga, trastornos en
la ubre como edemas y mastitis, desplazamiento del abomaso y
enfermedades podales. Conjuntamente con esta entidad morbosa, se
presentan otras patologías tales como fiebre de la leche,
cetosis, desplazamiento del abomaso, retención placentaria y
metritis (Álvarez, 1999a y Barros, 2001).
Cetosis.
Esta enfermedad ocurre frecuentemente en vacas de alta
producción en las primeras semanas después del parto
(González, 2000). La cetosis se desarrolla cuando la
gluconeogénesis no suple los requerimientos de glucosa del
animal, por lo que es necesaria la movilización de
ácidos grasos del tejido adiposo para suplir el déficit
energético con el consecuente aumento de la
cetogénesis. La escasez de oxaloacetato en su
doble papel, como precursor de la gluconeogénesis e
intermediario del ciclo de Krebs es de suma importancia en la
aparición de este trastorno. La carencia de oxaloacetato que
alimenta el ciclo de Krebs, proveniente del metabolismo de los
carbohidratos, hace que no
puede ser metabolizado por esta vía el acetil – CoA ruminal
(acetoacetato activado) o el producto de la Beta –
oxidación de los ácidos grasos, recurriendo a la
vía alternativa de la cetogénesis, generando un exceso
de cuerpos cetónicos (Mohar, 1992 y Alvarez, 1999a). La alta
concentración de proteínas en la dieta, asociada a la
ingestión de bajos niveles energéticos, se consideran
como los principales factores predisponentes a la aparición
del trastorno (Rajala – Schultz et al., 1999 y
González 2000).
Los principales síntomas se consideran la
pérdida del apetito, pérdida de peso, disminución
de la producción de leche y otros. Los principales signos metabólicos que
denotan su presencia están el aumento de los cuerpos
cetónicos y de los ácidos grasos libres en sangre,
hipoglicemia y disminución del glucógeno hepático
(Gonzáles, 2000). En estos casos la glicemia puede decaer de
2,8 – 3,9 mmol/L a niveles de 1,1 – 2,2 mmol/L, los
cuerpos cetónicos de niveles fisiológicos de 0,9
mmol/L, pueden ascender en sangre hasta valores de 9,6 mmol/L.
(Barros, 2001).
Síndrome de depresión de la grasa en
leche.
Como su nombre lo indica, se caracteriza por una
sensible disminución del contenido porcentual de grasa
láctea, que no está asociado con el inicio de la
lactación, sino a los regímenes alimentarios propios de
la producción intensiva de leche a base de dietas altas en
concentrados y bajas en alimentos fibrosos, los pastos, forraje y
heno. (Oldaham, 1991, Palmquist et al., 1993 y Pérez,
2001). La inclusión en la dieta de la vaca lechera de un
alto contenido de concentrados o cuando el forraje presente en la
dieta posee un tamaño de partícula menor a 3 mm, la
concentración de acético (AGV) en el rumen disminuye y
se incrementa en la misma proporción el contenido de
propiónico. La falta de acético provoca una carencia de
precursores para la síntesis de los ácidos grasos en la
glándula mamaria (Hurley, 2000).
Debido a que este trastorno provoca la distorsión
de la fermentación ruminal y un
grupo de desórdenes bioquímicos, la única medida
para su corrección es adecuar la relación
concentrado-fibra en la dieta, de tal forma, que se cubran los
requerimientos energéticos de la vaca y no se produzcan
alteraciones en el metabolismo ruminal negativas al metabolismo
general, ni alteraciones en la composición de la leche (Le
Ruyet y Tucker, 1992). También dietas altas en
proteínas son efectivas para prevenir la severidad de la
depresión de la grasa en la leche (Wittwer,
2000).
Hipocalcemia.
La también denominada fiebre de la leche o paresia
puerperal, se presenta entre el primer y tercer día
después del parto en vacas de alta producción. La vaca
presenta debilidad muscular, colapso circulatorio y
decúbito. Fredeen (1996) plantea que etiológicamente la
enfermedad manifiesta una disminución del calcio
plasmático, tanto el calcio total como el
ionizado.
Ello en gran medida se debe a un aporte inadecuado del
mismo en la dieta, justo cuando la demanda del mineral es
elevada. Cuantitativamente expresado, una vaca que produzca 30 Kg
de leche en un día a razón de 1,2 g de Ca/kg. de leche,
necesita 36 g de este macroelemento. Por otra parte, el calcio en
la leche es diez veces superior al calcio
plasmático.
De dicho calcio plasmático, es el calcio
iónico, (aproximadamente la mitad del calcio circulante), el
metabólicamente activo (González, 2000). A su vez la
hipocalcemia se puede presentar conjuntamente con trastornos del
equilibrio ácido – básico, aunque existen
reportes de predisposición genética, sobre todo que
dicho trastorno se presenta con mayor frecuencia en la raza
Jersey (Pérez, 2001). Otros cambios bioquímicos
concomitantes con la hipocalcemia son la hipofosfatemia,
hipermagnesemia, hiperglicemia e hipoinsulinemia. Se considera
que a niveles de calcio sanguíneo de 1,6 mmol/L comienzan a
observarse los primeros síntomas (Debilidad muscular, marcha
tambaleante y disminución de la producción
láctea). Sin embargo cuando el tenor de calcio
sanguíneo llega a ser menor de 1,1 mmol/L, la
sintomatología se hace manifiesta en su totalidad, cuando el
cuadro clínico se agrava, con peligro para la vida del
animal (Rajala – Schultz et al., 1999).
Hipomagnesemia.
Este trastorno metabólico es mundialmente conocido
como tetania de los pastos. Es típico de los países
templados con ganado estabulado durante el invierno. (White et
al., 2002). En este caso se presenta un síndrome
tetánico que ocurre al inicio de la primavera. Los
principales síntomas que se presentan dan como resultado
nerviosismo, temblor muscular, ataxia, espasmo, opistótonos
y convulsiones. La ocurrencia se asocia con una baja
disponibilidad de magnesio en la dieta o la baja capacidad de
absorber este mineral (Wittwer, 2000).
El tenor normal de magnesio en sangre que se encuentra
entre 0,8 a 1,2 mmol/L, decae a menos de 0,7 mmol/L durante la
aparición de esta patología. Los primeros síntomas
de irritabilidad aparecen cuando los niveles de este elemento
caen sobre 0,3 mmol/L y se agravan cuando disminuyen hasta 0,2
mmol/L (Barros, 2001).
Los efectos que sobre la composición de la leche y
sus propiedades provocan los trastornos metabólicos
descritos no son apreciables, pudiéndose señalar solo
disminuciones discretas en la grasa láctea. Las alteraciones
generalizadas en las propiedades físico-químicas de la
leche no han sido asociadas hasta el momento a estas
patologías (Ponce, 1999 y Ponce et al.,
2000).
Síndrome de Leche Anormal (SILA).
La disminución de los sólidos en la leche y al
alteración de sus propiedades físico-químicas es
un fenómeno recurrente que se ha profundizado en los
últimos años con grandes perjuicios para la industria
láctea, los productores y los consumidores en Cuba, al cual se le ha
denominado Síndrome de Leche Anormal (SILA).
El SILA se caracteriza por una depresión en los
sólidos de la leche, disminución de la capacidad
buferante, inestabilidad térmica de la leche y desequilibrio
mineral (Hernández y Ponce, 2005). Los indicadores más
sensibles del SILA son las bajas concentraciones de proteína
bruta y caseina (Menos de 2.90 y 2.20 g/% respectivamente),
disminución de la acidez titulable (Menor de 0.13 g/%) e
incremento del pH por encima de 6.75, así como
depresión en el contenido mineral, en especial de magnesio y
fósforo. Entre otras alteraciones que frecuentemente se
asocian están la disminución de la densidad (< 1.029
g/cm3) y trastornos en el tratamiento térmico de
la leche, así como reducción en los rendimientos y
calidad de los quesos.
Se ha definido que la causa fundamental del SILA se
asocia a la alimentación de rebaños lecheros altamente
especializados con caña de azúcar en los períodos
de seca y concomitante con una baja disponibilidad de pastos y
suministro inconsistente de forraje de calidad. El problemas se
acompaña con un importante desbalance
energía/proteína y que afecta la síntesis y
secreción láctea. La corrección de la dieta, la
implementación de un adecuado balance nutricional y la
utilización de formulaciones conteniendo sustancias
reguladoras y activadoras de la función ruminal permiten la
corrección del SILA (Hernández, 2003).
- Akers, R.M. (1990). Lactation physiology: A ruminant
animal perspective. Protoplasma 159:96-111. - Alvares, J. L.(1999)a. Bioquímica nutricional
y metabólica del bovino en el trópico. Ediciones
CENSA. - Alvares, J. L.(1999)b. Sistema Integral de
atención a la
Reproducción. Los conflictos entre la
reproducción y la producción de leche. Ediciones
CENSA. - Amiot, J. (1994). Ciencia y Tecnología de la
leche. Universite Laval Quebec. Edit Acribia, España. - Aranda, E.; Mendoza, G. D.; García –
Bojalil, C. y Castrejón, F. (2001). Growth of heifers
grazing stargrass complemented with sugar cane, urea and
protein supplement. Livestock Production Science 71:201
– 206. - Armenteros, Mabelin (1998). Evaluación de un
desinfectante mamario postordeño de origen natural.
Tesis en opción al grado científico de Doctor en
Ciencias. La Habana, Cuba. - Balbinotti, M.; Gonzales, H.L.; Fischer, V.; Ribeiro,
M.E.R.; Fagundes, C.M.; Schramm, R.; Stumpf, W. y Silva, M.A
(2001). Avaliacao dos sistemas de producao de leite de regiao
sul do Rio Grande do Sul e implicacoes na producao e qualidade
do leite. Arch. Latinoam. Prod. Animal. Vol.9,
suplemento 1 - Barbano, D.(1999): Trends in milk composition and
analysis in New York. In: Proceedings of the 1999 Cornell
Nutrition Conference for Feed Manufacturers. Rochester,
NY. - Barros, L.(2001): Trastornos metabólicos que
afetam a qualidade do leite. In: Uso do leite para monitorar
a nutricao e o metabolismo de vacas leiteiras. Gráfica
da Universidade Federal do Rio Grande do Sul, Porto
Alegre. - Beeyer D.E.; Rook A. J.; France J.; Dhanoa M.S. y
Gill, M. (1991). A review of empirical and mechanistic models
of lactational performance by the dairy cow. Livest. Prod.
Sci., 29, 115-130. - Bigner, D. R. ; Goff J. P.; Faust, M. A.;
Tyler, H. D. y Horst R. L.
(1997). Comparison of Oral Sodium Compounds
for the Correction of Acidosis.J. Dairy
Sci. 80:2162-2166. - Blackburn DG (1993). Lactation: Historical patterns
and potential for manipulation. J Dairy Sci
76:3195-3212 - Broderick, G. A. y Clayton, M. K.
(1997). A Statistical Evaluation of Animal and
Nutritional Factors Influencing Concentrations of Milk Urea
Nitrogen. J. Dairy Sci.
80:2964-2971. - Calvinho, L. (1995). La mastitis y su impacto en
la calidad de la leche. Informe Técnico INTA.
No. 1:1-14. - Cant, J. P.; Trout, D. R.; Qiao, F. y Purdie,
N. G. (2002). Milk Synthetic Response of the Bovine
Mammary Gland to an Increase in the Local Concentration of
Arterial Glucose. J. Dairy Sci.
85:494-503. - Clark, A.J. (1996) Genetic modification of milk
proteins. Am. J. Clin. Nutr. 63:633S-638S. - Contreras, P (2000): Indicadores do metabolismo
protéico utilizados nos perfis metabólicos de
rebanhos. In: Gonzales, F.H.; Barcellos, J.O.; Ospina, H.;
Ribeiro, L.A.: Perfil metabólico em ruminantes: seu uso
em nutricao e doencas nuricionais. Gráfica da
Universidade Federal do Rio Grande do Sul, Porto Alegre,
Brasil. - Coulon, J.B. y Pérochon, L. (2000). Evolution de
la production laitière au cours de la lactation :
modèle de prédiction chez la vache laitière
INRA Prod. Anim., 13, 349-360. - Dahl, G. E., Chastain, J. P. y Peters, R. R. (1998).
Manipulation of photoperiod to increase milk production in
cattle: biological, economic and practical considerations. pp.
259-265 In: Proceedings of the Fourth International Dairy
Housing Conference, J. P. Chastai, ed., Amer. Soc. Agric.
Engin., St. John's, MI - De Lima, Helenice; Fischer, Vivian; Ribeiro, Maria;
Medina, C.; Schrram, Renata y Stump, W.(2001). Variacao da
composicao do leite nos meses do ano sobre qualidade do leite.
Arch. Latinoam. Prod. Animal. Vol.9, suplemento
1. - Dils, R.R. (1986). Comparative aspects of milk fat
synthesis. J. Dairy Sci. 69:904-910. - Durr, J.W.; Fontaneli, R. S. y Burchard, J. F.
(2000). Fatores que afetam a composicao do leite. In: Curso
de sistemas de producao para gado de leite baseado em pastagens
sob plantio direto. Passo Fundo. Anais EMBRAPA –
Trigo. - Erdman, R. A. and M. Varner. (1995). Fixed yield
responses to increased milking frequency. J. Dairy Sci.
1199-1203. - Forsyth, I.A. (1989) Growth factors in mammary gland
function. J. Reprod. Fert. 85:759-770. - Fox, PF (1992) Advanced Dairy Chemistry,
Elsevier Applied Science, London - Fredeen, A. H. (1996): Considerations in the milk
nutritional modification of milk composition. Animal Feed
Science Technology 59:185-197. - Gonzáles, F.H.D (2000). Indicadores
sanguíneos do metabolismo mineral em ruminantes. In:
Gonzales, F.H.; Barcellos, J.O.; Ospina, H.; Ribeiro, L.A.:
Perfil metabólico em ruminantes: seu uso em nutricao e
doencas nuricionais. Gráfica da Universidade Federal
do Rio Grande do Sul, Porto Alegre, Brasil. - Gonzáles, F.H.D (2000).Uso de perfil
metabólico no diagnóstico de doencas metabólico
– ruminais em ruminantes. In: Gonzales, F.H.; Barcellos,
J.O.; Ospina, H.; Ribeiro, L.A.: Perfil metabólico em
ruminantes: seu uso em nutricao e doencas nuricionais.
Gráfica da Universidade Federal do Rio Grande do Sul,
Porto Alegre, Brasil. - Gonzáles, F.H.D.(2001): Composicao bioquimica
do leite e hormonios da lactacao. In: Uso do leite para
monitorar a nutricao e o metabolismo de vacas leiteiras.
Gráfica da Universidade Federal do Rio Grande do Sul,
Porto Alegre. - Gonzáles-Stagnaro, C.; Madrid-Bury, N. y Soto,
Belloso. E (1998). Mejora de la ganadería meztiza de
doble propósito. Facultad de Ciencias Veterinarias.
Ediciones Universidad del Zulia,
Venezuela. - Hernández, R. (2003): Caracterización,
diagnóstico y corrección de alteraciones en las
características físico-químicas de la leche en
rebaños Holstein Friesian. Tesis en opción al
Grado Científico de Doctor en Ciencias Veterinarias.
Universidad Agraria de La Habana, Cuba. - Hernández, R. y Ponce, P. (2005): Efecto de tres
tipos de dieta sobre la aparición de trastornos
metabólicos y su relación con alteraciones en la
composición de la leche en vacas Holstein Friesian.
Zootecnia. Trop. 23(3):295-310. ISSN
0798-7269. - Hof, G. M.; Vervoorn,D. P.; Lenaers, J. y
Tamminga S. (1997). Milk Urea Nitrogen as a Tool to
Monitor the Protein
Nutrition of Dairy Cows. J. Dairy
Sci. 80:3333-3340. - Hoffmeister D.F. (1989) Mammals of Illinois,
Univ. Illinois Press. - Hurley, W. L. (2000): Lactation biology.
University Press, University of Illinois. Urbana –
Champaing. - Hutjens, M.F.(1996): Milk Urea Nitrogen (MUN) as a
management tool. In: Ilinois Dairy Report, Building on
Basics, unviersity of Ilinois, Champaing, IL. - Ikonen, T. y Ojala, M. (1995). Effects of milk
protein genotypes on milk renneting properties assuming
alternative models. IDF Seminar “Implications of Genetic
Polymorphism of Milk Proteins on Production and Processing of
Milk, 28. – 29.3.1995 Zürich, Switzerland. Summary
2p. - Imagawa, W.; Yang, J.; Guzman, R. y Nandi, S. (1994)
Control of mammary gland development. Ch. 56 in The
Physiology of Lactation, 2nd edition, Eds. Knobil, K,
Neill, J., et al., Raven Press, NY, p. 1033. - Jensen, R.G.(1995): Handbook of milk
composition. Academic Press, San Diego. - Jonker,J. S.; Kohn, R. A. y Erdman, R. A.
(1998). Using Milk Urea Nitrogen to Predict Nitrogen
Excretion and Utilization Efficiency in Lactating Dairy Cows.
J. Dairy Sci.
81:2681-2692. - Juarez, F. I. ; Fox, D. G.; Blake R. W. y
Pell, A. N. (1999). Evaluation of Tropical Grasses for
Milk Production by Dual-Purpose Cows in Tropical Mexico.
J. Dairy Sci.
82:2136-2145. - Kalscheur, K. F.; Vandersall, J. H.; Erdman,
R. A.; Kohn R. A. y Russek-cohen E. (1999). Effects of
Dietary Crude Protein Concentration and Degradability on Milk
Production Responses of Early, Mid, and Late Lactation Dairy
Cows. J. Dairy Sci.
82:545-554. - Kaneko, J. J.,Harvey, J. W. y Bruss, M. L. (1997).
Clinical Biochemestry of Domestic Animals. San Diego,
Academic Press. - Kanno, C. (1990) Secretory membranes of the lactating
mammary gland. Protoplasma 159:184-208. - Kenelly, J (1999). Milk composition in the cow. In :
Proceedings of the 1999 Cornell Nutrition Conference for
Feed Manufacturers. Rochester, NY. - Klei, Linda R.; Lynch, Joanna M.; Barbano, D.
M.; Oltenacu, P. A.; Lednor, A. J. y Bandler, D. K.
(1997). Influence of Milking Three Times a Day on Milk
Quality. J. Dairy Sci.
80:427-436. - Knight, C.H. y Wilde, C.J. (1987) Mammary growth
during lactation: implications for increasing milk yield. J.
Dairy Sci. 70:1991-2000. - Lacy-Hulbert, S. J.; Woolford, M. W.;
Nicholas, G. D.; Prosser, C. G. y Stelwagen, K.
(1999). Effect of Milking Frequency and Pasture Intake
on Milk Yield and Composition of Late Lactation Cows.
J. Dairy Sci.
82:1232-1239. - Le Ruyet, P. y Tucker, W.B. (1992): Ruminal Buffers:
Temporal effects on buffering capacity and pH on ruminal fluid
from cows fed high concentrate diet. J. Dairy Sci. 75:
1069 – 1077. - Luquet, F.M. (1995).Leche y productos
lácteos. Edit. Acribia, España. - Mackle,T. R.; Bryant, A. M., Petch, S. F.,
Hill, J. P. y Auldist, M. J. (1999).
Nutritional Influences on the Composition of Milk from Cows of
Different Protein ghenotypes in New Zealand J. Dairy
Sci.
82:172-180. - Mao, I.L.; Buttazzoni, L.G. y
Aleandri, R. (1992). Effects of
polymorphic milk protein genes on milk yield and composition
traits in Holstein cattle. Acta Agric. Scand., Sect. A,
Animal Sci. 42:1-7. - Mepham, T.B. (1983): Biochemestry of
lactation. Elservier, London. - Mercier, J-C. y Vilotte, J-L. (1993) Structure and
function of milk protein genes. J. Dairy Sci.
76:3079-3098. - Mohar, F. (1992). Bioquímica Animal.
Ediciones ENPES, La Habana. - Montero, E. (2001). Situación actual y
perspectivas del mercado mundial de los
productos lácteos. In: Conferencia Internacional
"Aseguramiento de la Calidad en la Industria Láctea".
15 – 20 octubre, San José, Costa Rica. - National Research Council (2001). Nutrient
Requirements of Domestics Animals. National Academy of
Sciences, National Research Council. Sixth Revised Edition.
Washington DC. USA. - Ng-Kwai-Hang, K.F.; Hayes, J.F.; Moxley,
J.E. y Monardes,
H.G. (1984). Association of genetic variants
of casein and milk serum proteins in milk fat and protein
production by dairy cattle. J.Dairy Sci.
67:835. - Oldaham. J.D. (1991). Magnitud and implication of
changes in milk composition through manipulation of nutrition,
management and physiology. International Dairy Congress,
pág. 714-721. - Palmquist, D.L.; Beaulieu, A.D. y Barbano, D.M.
(1993). Feed and animal factors influencing milk fat
composition. J. Dairy Sci. 76:1753-1771. - Perez, J.R. (2001): O leite como ferramenta do
monitoramento nutricional. In: Uso do leite para monitorar a
nutricao e o metabolismo de vacas leiteiras. Gráfica
da Universidade Federal do Rio Grande do Sul, Porto Alegre.
Universidade Federal do Rio Grande do Sul, Porto
Alegre. - Pérochon, L. ; Coulon, J.B. y Lescourret,
F. (1996). Modelling lactation curves of dairy cows with
emphasis on individual variability. Anim. Sci., 63,
189-200. - Philpot , N (1992). Mastitis, el contrataque.
Edit. Philpot & Assoc. Inc. USA. - Place, J. y Gibson, P. (1988) Milk Composition.
Holstein Journal, August. - Pocknee B. (1998). Quality milk production – meeting
the challenge. Proceedings of the british mastitis
conference. Institute of animal health. P. 97 –
98. - Ponce, P (1999). Caracterizacao da Sindrome do leite
Anormal: Um enfoque das suas possiveis causas e correcao.
Anais do 4º Simposio Internacional sobre
Producao Intensiva de Leite, Caxambu, 22 – 24 de
julho. p. 61-76. - Ponce, P. (1984). Estudio de la lactancia en vacas de
los cruces 5/8 H – 3/8 C y ¾ H – ¼ C en
condiciones de Cuba. Tesis para la opción de candidato a
Doctor en Ciencias. La Habana. - Ponce, P. ; Hernández, R. ; Capdevila, J.;
Zaldivar, Victoria y Figueredo, J. (2000): Diagnóstico y
corrección de alteraciones en el contenido de solidos en
la leche asociados a trastornos metabólicos –
ruminales de la vaca lechera. Informe Final de Proyecto.
CENSA. - Ponce, P. y Bell, L (1984). Estudio de la lactancia
del cruce 5/8 H – 3/8 C cubano. I Componentes mayores de
la leche. Proteína, grasa, lactosa. Rev. Salud Anim.
6(1):85-94. - Ponce, P. y Bell, L (1985). Estudio de la lactancia
en vacas Holstein – Friesian, Cebú y sus cruces en
Cuba. Rev. Salud. Anim. 8(1):73-88. - Rajala-Schultz, P. J.y Gröhn, Y. T.;
Mcculloch, C. E. y Guard, C. L. (1999). Effects of
Clinical Mastitis on Milk Yield in Dairy Cows. J.
Dairy Sci.
82:1213-1220. - Rearte, D. (1993). Alimentación y
composición de la leche en los sistemas pastoriles.
Instituto Nacional de Tecnología Agropecuaria INTA.
Gráfica Lambertini, Argentina. - Sawyer, L. y Holt, C. (1993). The secondary structure
of milk proteins and their biological function. J. Dairy
Sci. 76:3062-3078. - Smith, K.L. (1996). El consejo Nacional de Mastitis y
el futuro del control de mastitis y la producción de leche
de calidad. Memorias del Congreso Regional del Consejo
Nacional de Mastitis de Estados Unidos de América.
Orlando, Colorado. - Strandberg E. y Lundberg C. (1991). A note on the
estimation of environmental effects on lactation curves.
Anim. Prod., 53, 399-402. - Tucker, H.A. (1994) Lactation and its hormonal
control. Ch. 57, In: The Physiology of Reproduction, 2nd
edition, Eds. E. Knobil, J. Neill, et al. Raven Press, Ltd.,
New York. p. 1065. - Van Soest, P.J., Robertson, J.B. y Lewis, B. A.
(1991). Symposium: Carbohidrate methodology, metabolism and
nutritional implications in dairy cattle. J. Dairy Sci.
74, 3583 – 3597. - Velmala, R., Mantysaari, E.
y Maki-Tanila, A.
(1993). Molecular genetic polymorphism at the
kappa-casein and beta-lactoglobulin loci in
Finnish dairy bulls. Agric.Sci.Finl.
2:431-435. - Walstra, P y Jenness, R (1984) Dairy Chemistry and
Physics, John Wiley & Sons, New York. - White, S.L.; Benson, G.A.; Washburn, S.P. y Green,
J.T. (2002). Milk production and economic measures in
confinement or pasture systems using seasonally calved Holstein
and Jersey cows. J Dairy Sci .85(1):95-104. - Wilde C.J. y Hurley, W.L. (1996) Animal models for
the study of milk secretion. J. Mammmary Gland Biol.
Neoplasia 1:123-134. - Wittwer, F. (2000). Diagnóstico dos equilibrios
metabólicos de energía em rebanhos bovinos. In:
Gonzales, F.H.; Barcellos, J.O.; Ospina, H.; Ribeiro, L.A.:
Perfil metabólico em ruminantes: seu uso em nutricao e
doencas nuricionais. Gráfica da Universidade Federal
do Rio Grande do Sul, Porto Alegre, Brasil.
Datos del autor.
Robier Hernández Rodríguez,
Doctor en Ciencias Veterinarias
Investigador Agregado
Centro Nacional de Sanidad Agropecuaria (CENSA)
San José de las Lajas
La Habana. Cuba