El Sistema de linfocitos T tiene restricción y si
la tiene, el linfocito T sólo es útil para el
individuo que la posee. Es por eso que en algunos experimentos, a
ratones de ciertas líneas puras se les ha sacado los
linfocitos T y se les han metido a otros ratones de otra
línea pura y no funcionan. A esto se le llama
restricción. Esto conlleva a decir que los linfocitos T
son ciegos inmunológicamente hablando, porque no reconocen
nada si el antígeno no es procesado y presentado.
Como todo lleva su jerarquía, las moléculas
más simples, más grandes (pero no complejas) llegan
hasta un cierto punto de reconocimiento. El linfocito T tiene a
su cargo las moléculas más complejas; él
procesa antígenos, pero además, es el
regulador.
Los antígenos pueden presentar diferente complejidad y
pueden ser de tres tipos:
- Timo independientes tipo I: aquellos antígenos
que son LPS puros de bacterias - Timo independientes tipo II: aquellos
antígenos con epitopos repetitivos lineales - Timo dependientes: su conformación es compleja
(digamos proteínas) o antígenos
monógenos grandes con muchas unidades repetitivas, pero
complejas
Ante la complejidad de los antígenos
timodependientes, el linfocito T requiere que el antígeno
le sea presentado y reconocido. Para eso se lo tienen que
presentar en el contexto de los Antígenos de
Histocompatilibidad (MHC), asociados a los Antígenos de
Histocompatibilidad. Por esta razón es que la respuesta de
los linfocitos T es restrictiva, porque los antígenos de
histocompatibilidad son homólogos pero son propios.
Existen diferentes células que van a tener la función de
presentar a el antígeno, de metabolizar el
antígeno. Generalmente son células
fagocíticas y entre ellas la principal es el (1)
macrófago. Todas las líneas de los
macrófagos principalmente son Células Presentadoras
de Antígenos (CPA), como unas (2) células
principales que están en los ganglios: células
dendríticas, (3) las que están debajo de la piel y
que van a ganglios: células de Langerhans y (4) los
linfocitos B.
Los inmunógenos a presentar pueden ser virus. Los virus,
como tienen unidades repetitivas y son complejas, son
inmunógenos complejos (timodependientes). Estos alertan,
en contacto con el receptor, a la célula
fagocítica. Se forma un fagosoma y luego éste se
une a un lisosoma, a pH
ácido, para degradar o metabolizar éste
inmunógeno (lo procesan). Luego los epitopos se unen a los
antígenos de histocompatibilidad. En este caso las
células linfoides principalmente secretan Antígenos
de Histocompatibildad clase II (MHC II) y entonces a el
antígeno lo asocian a moléculas de
histocompatibilidad clase II (asociación = epitopes + MHC
II) y luego lo presentan en la superficie. Al estar en la
superficie ya puede ser reconocido por un LINFOCITO T HELPER (ver
figura 3-2)
Los TCR de los linfocitos T helper son estructuras
que tienen funciones
similares a los receptores de los linfocitos T, que
también son receptores específicos que van a
activar clonalmente a determinado linfocito Th,
Esto sería principalmente una primera señal de la
activación del linfocito T helper. Pero aún
más, el macrófago o la Célula Presentadora
de Antígeno inmediatamente secreta una citocina para darle
la segunda señal de activación al linfocito T
helper y es la Interleucina I (IL I).
Entonces, las dos señales que nos van a activar a los
linfocitos T helper o ayudador son: (1) presentación del
antígeno con MHC II y (2) secreción de IL I (ver
figura 3-2). Con esto el linfocito comienza a activarse.
Importante: no todos los epitopes van a participar aquí y
no todos los linfocitos van a participar aquí; algunos
participan, los de la misma línea.
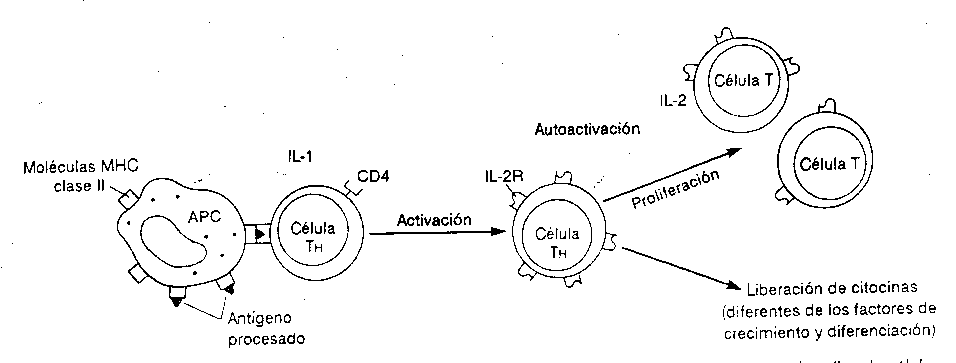
El linfocito T ya activado empieza a producir o
presentar en su superficie receptores para la siguientes
interleucinas, que es la Interleucina II (IL II) la cual ha sido
inducida por IL I. Entonces el linfocito Th secreta al medio la
IL II con el objetivo de
que ésta sirva como (1) amplificadora de la respuesta
inmune o de (2) crecimiento (esto quiere decir que como
ésta célula se ha activado se empieza a dividir
clonalmente). Entonces la IL II es un factor de
amplificación y un factor de crecimiento que forma miles y
miles de linfocitos T igual a ellos. Además, libera otras
citocinas: IL 3, Factor de Crecimiento de Células B
(BCGF), IL 6, TNF, Interfones, etc. y esto hace que actúen
otras células del sistema inmune con otras especificidades
y funciones. A partir de esto es que se afirma de que la
respuesta inmune específica es
heterogénea.

Ya tenemos activado el linfocito T. Su más
cercano colaborador a quien va a activar es al linfocito T
citotóxico. Una vez ya activado al linfocito T se procede
con la activación del linfocito T citotóxico, que
contribuye junto con el linfocito T en la erradicación de
la infección. Cuando el linfocito Th está activado,
secreta IL II y con ello está enviando una señal al
linfocito T CD8 para que éste se active. A eso se le
agrega que, como toda célula humana nucleada posee en su
superficie Moléculas de Histocompativilidad clase I (MHC
I), cuando una célula, ya sea que esté infectada
por virus, parásito o sea una célula tumoral,
presenta un antígeno en el contexto de MHC I y esto activa
al linfocito T citóxico (a ésta célula
humana se le llama célula en diana o célula diana,
célula blanco o a la que va dirigida la respuesta
inmune).
En conclusión, se tienen 2 señales para activar al
linfocito T citotóxico: (1) secreción de
interleucina clase II (IL II), y (2) presentación del
antígeno en el contexto de Antígeno de
Histocompatibilidad clase I (MHC I).
Al tener activado al linfocito T citotóxico, éste
se encarga de destruir , por medio de toxinas, ya sea
células infectadas intracelularmente (principalmente por
virus) o células tumorales.
Desde aquí, ya que se ha activado la respuesta inmune
celular (porque ésta es la que se ha activado), entra en
juego toda la
respuesta inmune: ya aquí en realidad entran los
linfocitos T a regular, magnificar o desacelerar ciertos
elementos o fenómenos del sistema inmune. Aquí el
linfocito se activaba sencillamente con unirse a una
molécula de inmunoglobulina en su superficie y con esto
basta para activarlo. Pero, estas moléculas
(antígenos) son timo independientes. Hay otros
antígenos que son complejos para el linfocito
B.
El linfocito B al entrar en contacto con
inmunógenos complejos, tiene capacidad no de realizar una
fagocitosis sino en realidad una endocitosis; al hacer esto, el
linfocito B procesa a dicho antígeno muy complejo y luego
lo presenta a linfocito T inductor, en el contexto siempre de
Antíegenos de Histocompatibilidad clase II (MHC II). Esto
lo hace porque el linfocito B está como "pidiendo el
auxilio" o "ayuda" al linfocito T.
Luego, el linfocito T le descarga 2 citocinas: (1) Factor de
Crecimiento de Células B (BCGF o "B-cell growth factor"),
para que esa célula se clone; (2) Factor de
Diferenciación de Células B (BCDF o "B-cell
differentiation factor"), para que el linfocito B se diferencie
en célula plasmática, que es la que principalmente
va a producir o secretar los Anticuerpos Específicos.
El linfocito B va a producir, en su progenie, tanto
células de memoria como células plasmáticas.
Las células de memoria probablemente se producen porque no
se alcanza, para una célula, la suficiente
concentración de Factor de diferenciación de
células B o BCDF y, en vez de transformarse a
células plasmáticas, se quedan como células
B de memoria.

Importante: Se dice también que las
células B y el linfocito T interactúan en la
Respuesta Inmune Secundaria, y que la Respuesta Inmune Primaria
es realizada principalmente por el macrófago. Entonces, es
la respuesta inmune secundaria la que está gobernada por
los linfocitos B o T.

Toda respuesta inmune específica está
gobernada por linfocito T helper o ayudador. Los colaboradores
del linfocito T son los macrófagos y los linfocitos T. De
aquí salen activados todos ellos y van a interactuar de
una forma directa o por medio de citocinas. Estas citocinas van a
activar a otras células como eosinófilos y,
además, van a ejercer acciones
quimiotácticas para neutrófilos, van a ir a ejercer
acciones desinfectorias de interferones, y otras citocinas van a
ejercer acciones de vasopermeabilidad para que lleguen los
linfocitos a ¿donde deben de estar?, etc. (ver
figura)
4.
Mecanismos de Eliminación de
Antígenos
- Neutralización de toxinas. Muchas
enfermedades, sobre todo bacterianas, dependen de la
liberación toxinas y las enfermedades son producidas
sobre todo por toxinas. Ej.: el Clostridium libera toxinas, el
cólera, difteria, tosferina, son enfermedades producidas
por toxinas. En estas enfermedades se producen anticuerpos
contra estas toxinas y se neutralizan mientras se lleva una
respuesta inmune celular más
específica. - Neutralización viral. El mecanismo principal
para combatir las infecciones virales es el destruir la
célula viralmente infectada, es una acción
citotóxica. Pero también se ha comprobado que hay
un fenómeno de neutralización viral, el cual se
puede explicar de la siguiente manera: muchas células de
nuestro cuerpo tienen receptores para algunos virus, la
neutralización viral se forma cuando un anticuerpo que
no permite la unión del virus con el receptor, por lo
que el virus no es capaz de infectar a la
célula.
- Opsonización de bacterias: aquí entran
algunos tipos de inmunoglogulinas, principalmente la Ig G y el
factor C3b del complemento, que mejoran la
fagocitosis. - Activación del complemento: la
activación completa de la cascada conlleva a una
citólisis de la célula - Citotoxicidad celular dependiente de anticuerpos
(ADCC): un anticuerpo es un coadyuvante de una célula
citotóxica, especialmente de la célula natural
asesina (NK), para destruir una célula. La principal
clase de anticuerpos, Ig G es la que se une a una subclase de
células NK (ver separata, "Mecanismos de
eliminación de antígeno")
 Página anterior Página anterior | Volver al principio del trabajo | Página siguiente  |