 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
Do nascer ao pôr-do-sol, foram registrados a cada 10 minutos a freqüência de bocados, peso de bocados, o teor de PB da forragem ingerida, o tempo de pastejo e o consumo de forragem. Para verificação da taxa de bocados, foi adotado o método do tempo de 20 bocados (Jamieson & Hodgson, 1979), realizando-se, posteriormente, a conversão dos valores obtidos para número de bocados por minuto. O peso de bocado foi determinado de forma simultânea à estimação da freqüência de bocado, por meio de coletas manuais realizadas de forma a reproduzir o peso de bocado de cinco ovelhas com mesmo peso e mesma largura de arcada dentária, escolhidas para este fim. Cada amostra destinada à determinação do peso de bocado foi composta por cinco coletas manuais (hand-plucking), cada uma equivalente a um bocado realizado pelo animal. A calibração da técnica de coleta manual foi feita de forma comparativa, utilizando-se uma ovelha fistulada no esôfago, a partir de uma regressão utilizada para corrigir os valores obtidos nas coletas manuais (Figura 1).

Utilizaram-se amostras coletadas através de fístula esofágica (constituídas pela forragem ingerida em 80 bocados) e por simulação manual de 20 bocados.
Posteriormente, as amostras obtidas pela coleta manual serviram também para determinação do teor de PB da forragem aparentemente ingerida.
O tempo de pastejo foi calculado a partir de registros da porcentagem de animais nesta atividade a cada 10 minutos, considerando todas as ovelhas do lote, pela seguinte fórmula:
TP10 min = 10*(% pastejo 1 + % pastejo 2 )/ 200,
em que: % pastejo 1 = % de animais pastejando em determinado horário; % pastejo 2 = % de animais pastejando no horário imediatamente posterior.
Desta forma, o tempo total de pastejo diurno correspondeu ao somatório dos valores calculados para cada intervalo de dez minutos.
O critério de observar todas as ovelhas do lote foi adotado pela maior praticidade encontrada pelos observadores de identificar determinada atividade em maiores distâncias sem a identificação individual dos animais, pois possuíam peso e condição corporal semelhantes, pertenciam à mesma raça e estavam em mesma fase fisiológica.
O consumo de forragem foi obtido pela seguinte fórmula:
Consumo = TP x FB x PB, em que: TP = tempo de pastejo; em que FB = freqüência de bocados; e PB = peso de bocados.
Os dados foram submetidos à análise de variância, em um delineamento inteiramente casualizado, em que as repetições consistiram dos cinco dias de observações do nascer ao pôr-do-sol, realizadas em cada estádio, EV, EP e EF (Pedroso et al., 2004a). Os tratamentos consistiram dos diferentes estádios fenológicos, de modo que, para a verificação do detalhamento do comportamento diurno, utilizou-se cada horário previamente definido como fator. Para as avaliações na pastagem, retiraram-se 26 amostras, marcadas por transectas atravessando diagonalmente o local, de modo a descrever um percurso em "W", visando à melhor representatividade da área. As amostras foram testadas quanto à significância da auto-correlação espacial e, na ausência de significância, foram consideradas repetições verdadeiras (Quadros & Bandinelli, 2005). Em todos os casos, as médias foram comparadas pelo teste de DMS com nível mínimo de 5% de significância.
A pastagem de azevém anual, no decorrer dos três estádios fenológicos, manteve-se com oferta média de folhas verdes superior a 5% do PV. Entretanto, em virtude da diminuição da participação de folhas na pastagem, com o avançar do ciclo de crescimento, a altura e a massa de forragem aumentaram para manutenção das ofertas pretendidas (Tabela 2). Desse modo, os animais apresentaram diferentes respostas comportamentais para cada estádio fenológico das plantas na pastagem.
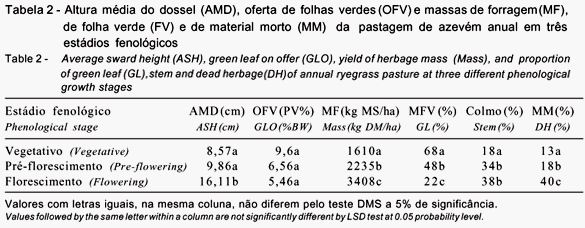
No estádio vegetativo, não foram verificadas variações significativas durante o período diurno para taxa de bocado, peso de bocado e teor de PB da forragem ingerida (Tabela 3). A alta participação de folhas verdes nesta fase provavelmente determinou valores máximos para estas variáveis comportamentais do nascer ao pôr-do-sol. Por outro lado, o tempo de pastejo variou com o avanço dos horários diurnos, resultando em variações no consumo de MS e PB pelos animais.

Os maiores valores (P<0,05) para tempo de pastejo e consumos de MS e PB foram verificados nos horários de 7 às 7h50 e de 16h às 18h e os menores, nos horários de 8h às 9h50. Considerando esses resultados, sugere-se que interferências na prática de pastejo para outros fins de manejo sejam feitas nos horários de 8h e 9h50, quando a atividade de pastejo é menor.
No estádio pré-florescimento, em decorrência do alongamento dos entrenós, a relação folha:colmo foi reduzida em comparação ao EV. Estas mudanças na estrutura e na qualidade da forragem ofertada determinaram mudanças comportamentais ao longo dos horários diurnos. Quando a maior parte do lote estava em pastejo, os animais estavam em intensa competição pelo consumo de forragem (das 7h às 7h50 e das 17h às 19h), verificando-se menor teor de PB na forragem ingerida, aumentos da freqüência de bocados e do peso de bocados e maiores consumos de MS e de PB (Tabela 4).

Essa estratégia reflete a tentativa do animal em garantir a quantidade de forragem necessária ao longo do dia, o que, provavelmente é conseqüência da maior competição pela forragem disponível, decorrente da maior presença de animais em atividade de pastejo em mesmo intervalo de tempo (Armstrong et al., 1995). Contudo, nos horários de menor atividade de pastejo, verificou-se maior intervalo de bocados e menor massa de forragem coletada por bocado, possibilitando a ingestão de frações com maior teor de PB (25,3%). Esse comportamento é considerado estratégia típica de ruminantes pela qual, depois de alcançada a saciedade física (resultante da estimulação dos receptores volumétricos do rúmen), inicia-se uma nova fase em que o aumento da qualidade da dieta é priorizado (Provenza & Lauchbaungh, 1999). Todavia, este comportamento refletiu-se em menores consumos de MS e PB em relação aos horários de maior intensidade de pastejo.
Os resultados sugeriram que práticas de manejo como a suplementação alimentar a campo devem ser feitas em horários próximos às 9h e ao meio-dia. Assim, seriam respeitados os principais horários de pastejo, em termos quantitativos (início da manhã e final de tarde), e os animais receberiam suplemento alimentar de melhor qualidade justamente nos horários em que são mais seletivos e que consomem menos. Rovira (1997), referindo-se ao comportamento ingestivo de bovinos, não recomendou a suplementação aos animais nas primeiras horas da manhã, pois, em vez de complementar, estar-se-ia substituindo a dieta disponível na pastagem pelo suplemento fornecido.
No estádio de florescimento, no entanto, houve certa uniformização da pastagem com a totalidade dos perfilhos em florescimento. Essa estrutura com elevada participação de colmo (38%) e material senescente (40%) provavelmente limitou a ação seletiva dos animais, que, conseqüentemente, apresentaram variações na taxa e no peso de bocados e no teor de PB ingerida durante o período diurno. É possível que o efeito conjunto da condição da pastagem e da elevação da temperatura neste estádio fenológico (EP - 15ºC; EF - 22ºC) tenha estimulado os animais ao pastejo durante o período noturno, na tentativa de satisfazer suas exigências nutricionais diárias, mesmo que não tenham sido alcançadas (Pedroso et al., 2004b). Parte desta explicação também reside no fato de que, nas primeiras horas do dia, registrou-se um curto tempo de atividade de pastejo (até as 8h50) e de baixos consumos de forragem e PB. Com a provável troca dos períodos de pastejo (preferência pelo pastejo noturno em vez de ao início do período diurno), houve maior necessidade de outras atividades como descanso e ruminação.
No restante do período diurno, o tempo de pastejo e, conseqüentemente, os consumos de MS e PB foram crescentes até o final da tarde (19h30). Estes dados sugerem que o momento mais adequado para intervenções no ritmo natural de pastejo seria nas primeiras horas do dia (até as 8h50), pois, nos primeiros horários matinais (7h às 7h50), a atividade de pastejo é menor.
Entretanto, nos três estádios fenológicos, ao final da tarde, foram verificados os maiores tempos de pastejo e consumos de MS e PB do período diurno. Vários autores registraram esses elevados valores momentos antes do anoitecer (Barbosa et al., 2004; Orr et al.,1997) e atribuíram o fato à composição química das plantas, que apresentam os maiores teores de MS e carboidratos nestes horários. Parsons et al. (1994) acrescentaram que a maior ingestão de forragem ao final da tarde ocorre porque os animais dispõem do período noturno para a atividade de ruminação.
Como resultado destas evoluções, foram verificados maiores consumos de MS e PB para EV e EP (Tabela 6). O consumo de PB foi muito próximo do recomendado para mantença de uma ovelha lactante com ganho médio diário de 25 g (240 g/dia). Todavia, o consumo de MS de 1.740 g/dia (Ganzábal, 1997), exigido por ovelhas em lactação, foi suprido em 62%.
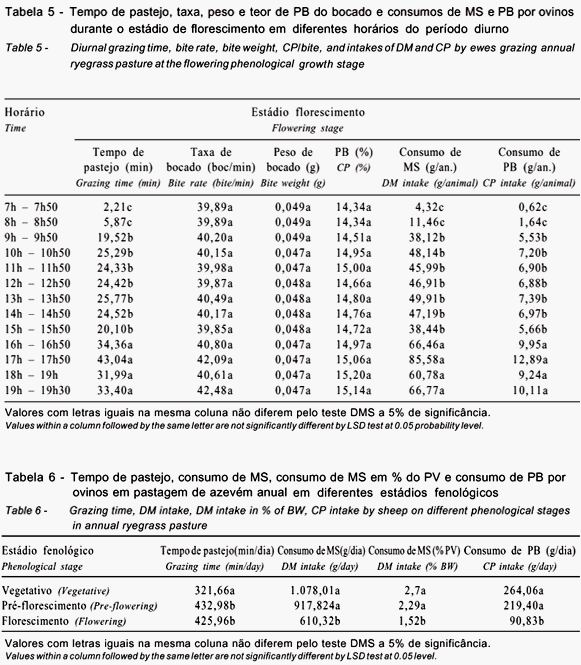
No estádio de florescimento, tanto o consumo diurno de MS como o de PB, foram considerados insatisfatórios para mantença destes animais de elevadas exigências nutricionais. De fato, o desempenho animal neste trabalho revelou uma importante perda de peso (152 g/dia) destes animais. Segundo Pedroso et al. (2004b), sob baixa qualidade, as variações de tempo de pastejo e no tamanho do bocado não permitem aos animais superar a dificuldade imposta pela condição da pastagem em florescimento para colher uma dieta que atenda às suas exigências nutricionais para mantença e aleitamento.
Os maiores tempos de pastejo e consumos de MS e PB ocorrem no final do período diurno para os estádios vegetativo, pré-florescimento e de florescimento.
Nos dois estádios iniciais, estes maiores valores também ocorrem nos primeiros momentos de luz natural.
No pré-florescimento, em alta atividade de pastejo, as ovelhas elevam a taxa de bocados e a quantidade de forragem coletada por bocado e diminuem a qualidade do ingerido. No estádio de florescimento, reduzem os tempos de pastejo e o consumo de MS e de PB ao longo de todo o período diurno, como conseqüência da baixa qualidade da forragem disponível e da estrutura da vegetação.
ARMSTRONG, R.H.; ROBERTSON, E.; HUNTER, E.A. The effect of sward height and its direction of change on the herbage intake, diet selection and performance of weaned lambs grazing ryegrass swards. Grass and Forage Science, v.50, p.389-398, 1995.
BARBOSA, C.M.P.; CARVALHO, P.C.F.; SILVA, M.A. et al. Comportamento ingestivo diurno de cordeiros em pastagem de azevém anual. In: REUNIÃO ANUAL DA SOCIEDADE BRASILEIRA DE ZOOTECNIA, 41., 2004, Recife. Anais... Recife: Sociedade Brasileira de Zootecnia, 2004. (CD-ROM).
BERGGREN-THOMAS, B.; HOHENBOKEN, W.D. The effects of size-breed, forage availability and weather on the grazing behaviour of crossbred ewes. Applied Animal Behavior Science, v.15, p.217-228, 1986.
EMPRESA BRASILEIRA DE PESQUISA E AGROPECUÁRIA - EMBRAPA. Sistema Brasileiro de Classificação de Solo. Brasília: Embrapa Cerrados, Rio de Janeiro: Centro Nacional de Pesquisa de Solos, 1999. 412p.
GANZÁBAL, A. Alimentación de ovinos com pasturas sembradas. Montevideo: Instituto Nacional de Investigación Agropecuaria, 1997. v.84, 44p.
JAMIESON, W.S.; HODGSON, J. The effect of daily herbage allowance and sward characteristics upon the ingestive behaviour and herbage intake of calves under strip-grazing for grazing dairy cows. Grass and Forage Science, v.34, p.69-77, 1979.
MOORE, K.J.; MOSER, L.E.; VOGEL, K.P. et al. Describing and quantifying growth stages of perennial forage grasses. Agronomy Journal, v.83, p.1073-1077, 1991.
ORR, R.J.; PENNING, P.D.; HARVEY, A.; CHAMPION, R.A. Diurnal patterns of intake rate by sheep grazing monocultures of ryegrass or white clover. Applied Animal Behavior Science, v.52, p.65-77, 1997.
PARSONS, A.J.; NEWMAN, J.A.; PENNING, P.D. et al. Diet preference of sheep: effects of recent diet, physiological state and species abundance. Journal of Animal Ecology, v.63, p.465-478, 1994.
PEDROSO, C.E.S.; MEDEIROS, R.B.; ABREU DA SILVA, M. et al. Comportamento de ovinos em gestação e lactação sob pastejo em diferentes estádios fenológicos de azevém anual. Revista Brasileira de Zootecnia, v.33, n.5, p.1340-1344, 2004a.
PEDROSO, C.E.S.; MEDEIROS, R.B.; ABREU DA SILVA, M. et al. Produção de ovinos em gestação e lactação sob pastejo em diferentes estádios fenológicos de azevém anual. Revista Brasileira de Zootecnia, v.33, n.5, p.1345-1350, 2004b.
PONTES, L.S.; NABINGER, C.; CARVALHO, P.C.F. et al. Variáveis morfogênicas e estruturais de azevém anual (Lolium multiflorum Lam.) manejado em diferentes alturas. Revista Brasileira de Zootecnia, v.32, n.4, p.814-820, 2003.
PRACHE, S. Intake rate, intake per bite and time per bite of lactating ewes on vegetative and reproductive swards. Applied Animal Behavior Science, v.52, p.53-64, 1997.
PROVENZA, F.D.; LAUCHBAUNGH, K.L. Foraging on the edge of chaos. In: LAUCHBAUNGH, K.L.; MOSLEY, J.C.; SANDERS, K.D. (Eds.). Grazing behavior of livestock and wildlife. Moscow: University of Idaho, 1999. p.1-12.
QUADROS, F.L.F.; BANDINELLI, D.G. Efeitos da adubação nitrogenada e de sistemas de manejo sobre a morfogênese de Lolium multiflorum Lam. e Paspalum urvillei Steud. em ambiente de várzea. Revista Brasileira de Zootecnia, v.34, n.1, p.44-53, 2005.
ROOK, A.J.; HARVEY, A.; PARSONS, A.J. et al. Effect of long term changes in relative resource availability on dietary preference of grazing sheep for perennial ryegrass and white clover. Grass and Forage Science, v.57, p.54-60, 2002.
ROVIRA, J. Manejo nutritivo de los rodeos de cria en pastoreo. Montevideo: Hemisferio Sur, 1997. 288p.
SOARES, A.B.; MEZZALIRA, J.C.; BUENO, E.A.C. et al. Efeitos de diferentes intensidades de pastejo em pastagem nativa melhorada sobre o desempenho animal. Revista Brasileira de Zootecnia, v.35, n.1, p.75-83, 2006.
THOMAS, B.B.; HOHENBOKEN, W.D. The effects of size-breed, forage availability and weather on the grazing behaviour of crossbred ewes. Applied Animal Behavior Science, v.15, p.217-228, 1986.
Renato Borges de MedeirosI;
Carlos Eduardo da Silva PedrosoII;
João Batista Jornada da JornadaII;
Marcelo Abreu da SilvaI;
João Carlos de SaibroIII
IDPFA - Faculdade de Agronomia, UFRGS, Porto Alegre, RS, CEP: 91540-000
IIMestrando - DPFA, Fac. de Agronomia, UFRGS
IIIColaborador Convidado, DPFA - Faculdade de Agronomia, UFRGS
Revista Brasileira de Zootecnia v.36 n.1 Viçosa jan./fev. 2007
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




