 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
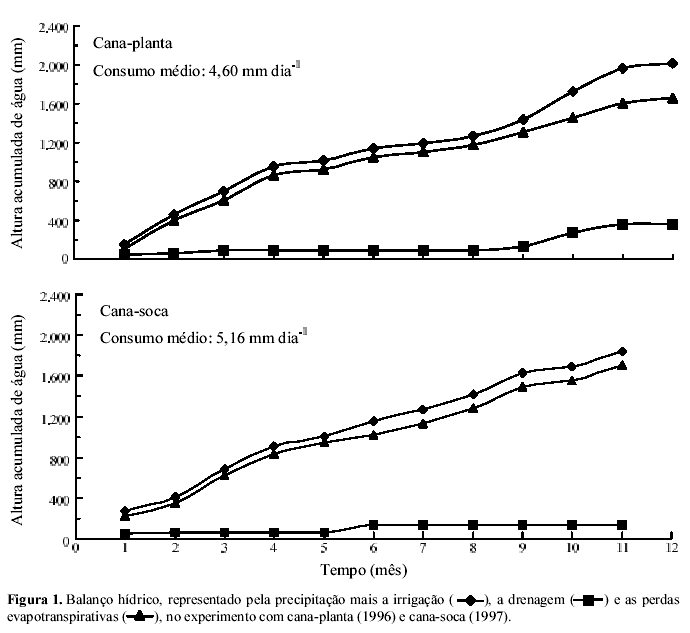
Os estudos foram desenvolvidos no campo experimental do posto meteorológico do Departamento de Ciências Exatas da Esalq/USP, em Piracicaba, SP. O experimento com cana-planta se estendeu de janeiro a dezembro de 1996. O volume total de água recebido pela cultura, de precipitações e de irrigações, nas épocas de estiagem, foi de 2.014 mm (1.255 e 759 mm, respectivamente, chuvas e irrigação), e as médias das temperaturas máximas e mínimas foram de 28,7 e 15,8°C, respectivamente. No decorrer do segundo experimento, de janeiro a novembro de 1997, as médias das temperaturas máximas e mínimas foram de 28,4 e 15,5°C, respectivamente. A cultura recebeu uma lâmina de água de 1.839 mm, sendo 923 mm de precipitações e 916 mm de irrigações.
O experimento constou de 48 parcelas constituídas por vasos de 220 L (60 cm de diâmetro e 90 cm de altura) contendo terra de textura arenosa (840, 60 e 100 g kg-1, respectivamente, de areia total, silte e argila) com as características químicas: pH, 4,4 (CaCl2); 19 g kg-1 de MO; 10 mg dm-3 de P (resina); K, Ca, Mg e H+Al: 1, 17, 3 e 34 mmolc dm-3, respectivamente. Os vasos possuíam, na parte inferior, um dreno constituído de uma camada de 10 cm de brita zero, coberta por manta Bidim, e foram mantidos ao ar livre.
O experimento com cana-planta (24 vasos), instalado em janeiro de 1996, constou de dois tipos de restos culturais incorporados ao solo, simulando a reforma de canavial após a colheita mecânica da canadeaçúcar, com queima (CQ) ou sem prévia despalha a fogo (SQ); e de quatro doses de N no plantio: 0, 30, 60 e 90 kg ha-1. No SQ, os restos culturais ¾ folhas secas (177 g por vaso de matéria seca (MS); 386 e 3,99 g kg-1, de C e N, respectivamente), colmos aéreos e ponteiros (122 g por vaso de MS; 423 e 7,84 g kg-1 de C e N, respectivamente), e rizomas (251 g por vaso de MS; 400 e 5,70 g kg-1 de C e N, respectivamente) ¾ foram misturados à terra dos vasos (025 cm de profundidade) em quantidades equivalentes a 6,3, 4,3 e 8,9 Mg ha-1 de matéria seca (MS total equivalente a 19,5 Mg ha-1), respectivamente, referidas com base em superfície e considerada a área dos vasos como sendo 0,283 m2. Esses valores foram definidos tomando por base as produções médias de resíduos culturais obtidas por Camargo (1989), Abramo Filho et al. (1993) e Trivelin et al. (1995). No CQ os restos culturais foram misturados ao solo nas mesmas quantidades acima especificadas, excluindo-se as folhas secas, procurando-se, assim, simular uma condição de reforma do canavial com despalha a fogo antes da colheita mecanizada; a quantidade equivalente de restos culturais foi de 13,2 Mg ha-1. Todos os restos culturais foram grosseiramente triturados antes de misturados ao solo. O experimento com cana-soca (24 vasos) constou da implantação, em janeiro de 1996, também dos tratamentos SQ e CQ.
Na camada superficial da terra de todos os 48 vasos (025 cm de profundidade e correspondente a 90 kg de terra), adicionaram-se os restos culturais, conforme o tratamento, e CaO e MgO, numa relação 2:1 (m/m), em quantidade suficiente para elevar a 60% a saturação por bases. Dez dias após a adição do corretivo e dos restos culturais ao solo, realizou-se a adubação de plantio. Transplantaram-se três mudas de cana-de-açúcar da cultivar SP 80-1842, com, aproximadamente, 20 cm de altura, para cada vaso. A adubação de plantio foi realizada no fundo de cova com 15 cm de profundidade. O KCl e o superfosfato triplo foram aplicados em todas as parcelas, na dose equivalente a 120 kg ha-1 dos respectivos óxidos. Nas parcelas do experimento com cana-planta aplicaram-se 0, 899,2, 1.798,4 e 2.697,6 mg de N-uréia por vaso, com 10,10% de átomos de abundância de 15N, equivalentes a 0, 30, 60 e 90 kg ha-1 de N. As parcelas destinadas ao experimento com cana-soca receberam 899,2 mg de N por vaso (uréia), equivalente à dose de 30 kg ha-1 de N.
O experimento com cana-planta (24 parcelas) foi colhido em dezembro de 1996. Na mesma época, fez-se o primeiro corte da parte aérea das plantas das parcelas com cana-soca, instalando-se os tratamentos desse experimento. Nas parcelas do tratamento SQ, a superfície do solo foi coberta com palha de cana-de-açúcar, constituída por folhas secas e ponteiros grosseiramente triturados; no CQ, o solo ficou descoberto, aplicando-se na superfície as cinzas resultantes da queima de quantidade correspondente de palha do tratamento SQ. A massa de matéria seca de palha colocada por vaso foi, em média, de 592 g, equivalente a 21 Mg ha-1.
No experimento com cana-soca, estudaram-se duas formas de aplicação da uréia nos tratamentos SQ e CQ: em superfície, sobre a palhada ou sobre o solo descoberto, e na profundidade de 15 cm, e duas fontes de potássio: KCl ou vinhaça. Utilizou-se solução de uréia (78,4 g L-1) com 10,08% de átomos de 15N. As quantidades de N e K2O aplicadas por vaso foram, respectivamente, de 2.643 e 3.510 mg, e equivalentes a 90 e 120 kg ha-1. Esse experimento foi colhido em novembro de 1997.
Tanto no experimento com a cana-planta (1996) como no de cana-soca (1997), foi sempre recolhida a solução percolada no solo, que atingia o dreno em cada vaso, sendo nela determinado o N mineral total: N-NH4+ e N-(NO3-+NO2- ), conforme Keeney & Nelson (1982), e de abundância de 15N, por espectrometria de massas (Trivelin et al., 1973).
Ao final de cada experimento efetuou-se a colheita das plantas, amostrando-se a parte aérea e a subterrânea, e do solo de todas as parcelas, após a retirada da parte subterrânea da cana-de-açúcar. No segundo experimento, também foi amostrada a palhada residual na superfície do solo. Procederam-se as determinações de N-total e de abundância de 15N em amostras de plantas, de solo e, também, da palhada residual no segundo experimento (Trivelin et al., 1973).
A recuperação porcentual do N-uréia (R) foi determinada pela expressão:
![]()
sendo:
![]()
onde: A e B abundância de 15N (% de átomos) da planta e da uréia-15N, respectivamente; C a abundância natural de 15N (0,366% de átomos); NT é o conteúdo de nitrogênio da planta (grama por parcela) e Naf a quantidade de N-uréia aplicada (miligrama por parcela).
O delineamento experimental foi o de blocos ao acaso, com três repetições, e os resultados de recuperação do N-uréia foram submetidos à análise da variância (teste F a 95% de probabilidade), sendo o teste de Tukey (p = 0,05) usado nas comparações de médias dos tratamentos.
Houve efeito de resíduos culturais na recuperação do N da uréia no solo no experimento com cana-planta, evidenciando no tratamento que simulou a reforma de canavial sem queima (SQ) a maior média (Tabela 1). Esse resultado indica haver uma relação direta entre a quantidade de resíduos culturais incorporados e o efeito residual do N-uréia no solo. Nas condições do experimento, não ocorreu na planta efeito dos fatores estudados na recuperação porcentual do Nfertilizante. Durante o período experimental, também não houve perdas mensuráveis de N derivado da uréia (15N) por lixiviação, na profundidade de 90 cm. O N total lixiviado foi equivalente a 4,5 kg ha-1 de N e oriundo do N mineralizado no solo (Oliveira et al., 1999).
Uma vez que foram tomados cuidados para evitar erros sistemáticos, as perdas se deram do sistema solo-planta para a atmosfera (Tabela 1). Essas perdas podem ter ocorrido, no todo ou em parte, por desnitrificação do NO3 derivado da uréia. No experimento existiram condições favoráveis à desnitrificação. A incorporação dos restos culturais ao solo e o consumo de oxigênio pelos microrganismos mineralizadores dessa matéria orgânica resultaram, muito provavelmente, em condição anóxica no solo, levando à reação de redução do NO3 pelos microrganismos desnitrificadores, quimiorganotróficos anaeróbios facultativos. Além disso, as chuvas ocorridas no início e no fim do experimento mantiveram o solo saturado com água, favorecendo a anaerobiose (Figura 1). Dobbie et al. (1999) encontraram perdas por desnitrificação em culturas como cana-de-açúcar fertilizada com N e irrigada, pastagem e florestas tropicais, variáveis de 0,3 a 18,4 kg ha-1 de N-N2O, e verificaram que elas cresciam exponencialmente com o preenchimento do espaço poroso do solo pela água. Outra possibilidade que justificaria as perdas (Tabela 1) refere-se à volatilização de amônia pela parte aérea da cultura. Entre as duas, nesse caso, a desnitrificação parece ser a mais provável.
Em relação às trocas de amônia que normalmente ocorrem entre os vegetais e a atmosfera, é possível inferir que a cana-de-açúcar, no ciclo de cana-planta, considerando-se neste experimento a cultivar SP 801842, com características de precocidade (Quarta..., 1993), aliada às condições experimentais e fatores meteorológicos, em especial às hídricas, apresentou ponto de compensação para amônia inferior à concentração de NH3 da atmosfera, o que favoreceu a absorção, sem que houvesse emissão de amônia. Essa hipótese se apóia na significativa participação no conteúdo de N da cana-planta de outras fontes de N e no N absorvido da uréia que representou, em média, 11,5% do acumulado na planta toda (7,0%, 11,7% e 15,9%, respectivamente, para as doses de N equivalentes a 30, 60 e 90 kg ha-1) (Trivelin, 2000), e a melhor eficiência que a canaplanta apresenta comparativamente às soqueiras, em relação ao uso do N, ou seja, quilograma de N utilizado por megagrama de colmos produzidos (Silva & Casagrande, 1983). Ainda, nesse mesmo contexto, considerando as perdas verificadas neste e em trabalhos precedentes (Camargo, 1989; Trivelin et al., 1996), poder-se-á estender a hipótese formulada, inferindo-se que as rebrotas de cana-de-açúcar, dependendo das condições meteorológicas, apresentariam maior ponto de compensação para amônia, o que justificaria as perdas de N por volatilização pela folhagem da cultura.
No ciclo de cana-soca, somente o fator localização do fertilizante teve efeito na recuperação do N da uréia, no solo a uréia aplicada em superfície mostrou menor efeito residual do que a aplicada em profundidade (Tabela 2), provavelmente, em vista das maiores perdas de amônia por volatilização (Trivelin et al., 1994). Na palha, o maior efeito residual do N-uréia ocorreu nos tratamentos em que o fertilizante foi aplicado em superfície, indicando que houve imobilização do N-fertilizante pela biomassa microbiana, que se desenvolveu nesse material de alta relação C:N. Os resultados de imobilização do N da uréia aplicada em superfície foram superiores aos de Gava (1999), que obteve recuperação de 8±2%. No presente trabalho, a uréia foi aplicada como solução concentrada sobre a palhada, após a aplicação da vinhaça, enquanto Gava (1999) aplicou mistura de vinhaça e uréia (100 m3 ha-1 de vinhaça e 100 kg ha-1 de Nuréia), o que pode ter facilitado a penetração da uréia no solo, reduzindo a imobilização pelos microrganismos na palhada. Por sua vez, a presença, na palha, do N da uréia aplicada em profundidade (Tabela 2) pode ser devido ao movimento ascendente, no solo, desse N, que foi imobilizado pelos microrganismos, ou, o que é mais plausível, que as raízes da soca que se desenvolveram na superfície, logo abaixo da palhada, beneficiadas pelas condições microclimáticas da cobertura do solo, fizeram parte da amostra de palha, causando o enriquecimento em 15N desse material.
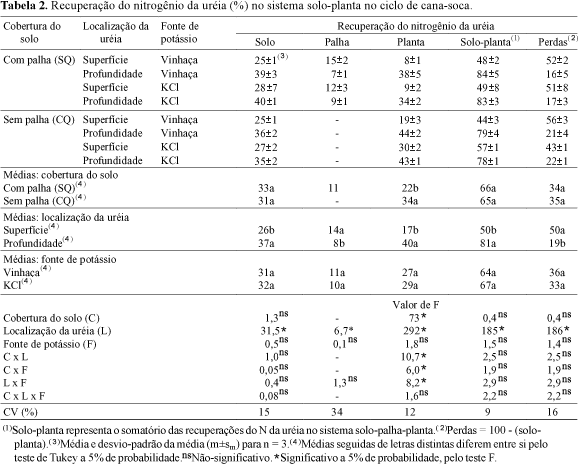
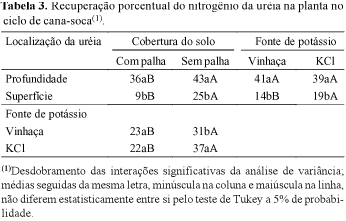
Na recuperação do N-uréia na planta toda (parte aérea e sistema radicular da cana-soca), houve efeito da localização da uréia, da cobertura do solo e, também, de interações entre os fatores de estudo (Tabela 2). Com a aplicação do fertilizante em profundidade, no solo, a cana-soca recuperou, em média, 40% do N-uréia, representando mais do dobro do adubo aplicado em superfície. Esse resultado foi inferior à recuperação pela cana-planta (54%) no primeiro experimento (Tabela 1), e equivalente à obtida por Trivelin et al. (1995), em relação à parte aérea de cana-soca de final de safra, em condições de campo, com uréia aplicada em profundidade. Na condição de solo coberto com palha (SQ), a recuperação do N-uréia na cana-soca foi inferior à sem palha (CQ). Os resultados do desmembramento das interações significativas da análise de variância indicaram de forma geral, independentemente da fonte de K, que as maiores recuperações na planta ocorreram sem a cobertura de palha e com a uréia aplicada em profundidade no solo (Tabelas 2 e 3). Esses resultados evidenciaram que a aplicação da uréia em superfície e em solo coberto com palha, resultaram nas menores recuperações do N-fertilizante na cana-soca, decorrentes, em grande parte, de perdas do Nuréia por volatilização de amônia, no solo.
Somente o fator localização do fertilizante mostrou efeito na recuperação do Nuréia no sistema solo-planta (Tabela 2). Sua aplicação em profundidade no solo resultou em 81% de recuperação, enquanto a superficial, somente em 50%. A cobertura do solo, simulando as condições com e sem queima do canavial (CQ e SQ), e a fonte de K não mostraram efeito na recuperação do N da uréia no sistema solo-planta. Esses resultados indicam que no manejo de canaviais sem ou com queima ¾ com ou sem palha cobrindo a superfície do solo ¾ a aplicação da uréia deve, preferencialmente, ser feita em profundidade.
Considerando-se que neste experimento, também não se constatou a lixiviação do N-fertilizante (Oliveira, 1999), as perdas do sistema solo-planta, no tratamento com fertilizante aplicado em profundidade, representaram as perdas para a atmosfera (Tabela 2). As perdas de 50% do N-uréia aplicado em superfície representam aquelas que ocorreram no solo, principalmente, por volatilização de amônia, e pela parte aérea da cultura.
No experimento com cana-soca, nas condições de solo coberto, ou não, com palha, não deve ter ocorrido a estimulação de atividade microbiana, pelo menos com a intensidade que se supôs no experimento com cana-planta e, conseqüentemente, não deve ter havido consumo excessivo de oxigênio do ar do solo, caracterizando condições extremas de anaerobiose que justificassem a desnitrificação. Durante a realização deste experimento, ao contrário do primeiro, a pluviosidade foi baixa (923 mm) o que exigiu a aplicação de 916 mm de água, ficando reduzida a possibilidade da desnitrificação. Assim, pode-se afirmar que, nos tratamentos com uréia aplicada em superfície, as perdas deveram-se à volatilização de amônia do solo e da folhagem. Nos tratamentos com aplicação da uréia em profundidade, o resultado médio de 19% representa as perdas pela parte aérea das plantas, uma vez que é desprezível a volatilização de amônia do solo (Tabela 2).
Os resultados de perdas do N-fertilizante do sistema soloplanta, nos dois experimentos, parecem indicar que o ponto de compensação para absorção/emissão de amônia na cana-de-açúcar não foi o mesmo nos ciclos de cana-planta e cana-soca. Na cana-planta, o índice pode ter sido inferior à concentração de NH3 da atmosfera, prevalecendo a absorção, enquanto, na rebrota, o ponto de compensação para amônia foi superior à concentração de amônia do ar, pelo menos no período compreendido entre a metade e o fim do ciclo da cultura, quando ocorrem as perdas, como constatado indiretamente por Ng Kee Kwong & Deville (1994a, 1994b) e Trivelin et al. (1996). Uma possível explicação para esses resultados pode estar no regime hídrico (Figura 1), associado aos fatores meteorológicos como temperatura do ar, radiação solar e velocidade do vento, que levou a cana-soca a apresentar maior perda de N pela parte aérea, com aumento no ponto de compensação de amônia. Plantas em condições de deficiência hídrica aceleram a degradação de proteínas, diminuindo a síntese (Hasiao, 1973; Du et al., 1996), que, por conseqüência, pode levar a perdas de N pela parte aérea.
Supondo-se que no caso de as perdas do N-uréia em cana-soca, com o fertilizante aplicado em profundidade, terem ocorrido unicamente por volatilização de NH3 da parte aérea, e considerado, ao final do ciclo, um acúmulo de N na parte aérea da cultura de aproximadamente 200 kg ha-1, sendo 20% desse N, como valor médio (Trivelin et al., 1995, 1996), derivado da uréia, as perdas totais (N derivado da uréia e de outras fontes assimilado pela cana-de-açúcar) foram da ordem de 90 kg ha-1 de N.
1. As perdas do N-uréia no sistema solo-planta no ciclo de cana-soca são superiores às de cana-planta.
2. No ciclo de cana-soca, as maiores perdas do Nuréia no sistema solo-planta ocorrem quando o fertilizante é aplicado na superfície.
3. As perdas de N pela folhagem da cana-de-açúcar, no ciclo de cana-soca, são da ordem de 90 kg ha-1 de N.
À Usina Costa Pinto S.A. Açúcar e Álcool e ao Centro de Tecnologia da Copersucar, Divisão de Agronomia e Engenharia Agrícola, pelo apoio na execução deste estudo; à Fapesp e CNPq, pelo financiamento da pesquisa.
ABRAMO FILHO, J.; MATSUOKA, S.; SPERANDIO, M. L.; RODRIGUES, R. C. D.; MARCHETTI, L. L. Resíduo da colheita mecanizada de cana crua. Álcool & Açúcar, São Paulo, n. 67, p. 23-25, abr./maio 1993.
BOUWMAN, A. F.; LEE, D. S.; WAH, A.; DENTENER, F. .J.; VANDERHOEK, K. W.; OLIVIER, J. G. J. A global high-resolution emission inventory for ammonia. Global Biogeochemical Cycles, Washington, v. 11, n. 4, p. 561-587, 1997.
CAMARGO, P. B. Dinâmica do nitrogênio dos fertilizantes uréia (15N) e aquamônia (15N) incorporados ao solo na cultura de cana-de-açúcar. Piracicaba: Esalq, 1989. 104 p. Dissertação de Mestrado.
DOBBIE, K. E.; McTAGGART, I. P.; SMITH, K. A. Nitrous oxide emissions from intensive agricultural systems: variations between crops and seasons, key driving variables, and mean emission factors. Journal of Geophysical Research Atmosphere, Washington, v. 104, n. D21, p. 26891-26899, 1999.
DU, Y. C.; KAWAMITSU, Y.; NOSE, A.; HIYANE, S.; MURAYAMA, S.; WASANO, K.; UCHIDA, Y. Effects of water stress on carbon exchange rate and activities of photosynthetic enzymes in leaves of sugarcane (Saccharum sp.). Australian Journal of Plant Physiology, Collingwood, v. 23, n. 6, p. 719-726, 1996.
DUXBURY, J. M. The significance of agricultural sources of greenhouse gases. Fertilizer Research, Dordrecht, v. 38, n. 1, p. 151-163, 1994.
FARQUHAR, G. D.; FIRTH, P. M.; WETSELAAR, R.; WEIR, B. On the gaseous exchange of ammonia between leaves and the environment: determination of the ammonia compensation point. Plant Physiology, Rockville, v. 66, p. 710-714, 1980.
GAVA, G. J. C. Utilização do nitrogênio da uréia (15N) e da palhada (15N) por soqueira de cana-de-açúcar no manejo sem despalha a fogo. Piracicaba: USP, 1999. 81 p. Dissertação de Mestrado.
HASIAO, T. C. Plant response to water stress. Annual Review of Plant Physiology, Palo Alto, v. 24, p. 519-570, 1973.
HOLTAN-HARTWIG, L.; BOCKMAN, O. C. Ammonia exchange between crops and air. Norwegian Journal of Agricultural Sciences, Aas, v. 14, p. 1-41, 1994. Supplement.
HUSTED, S.; MATTSSON, M.; SCHJOERRING, J. K. Ammonia compensation points in two cultivars of Hordeum vulgare L. during vegetative and generative growth. Plant Cell and Environment, Oxford, v. 19, n. 11, p. 1299-1306, 1996.
KEENEY, D. R.; NELSON, D. W. Nitrogen: inorganic forms. In: PAGE, A. L.; MILLER, R. H.; KEENEY, D. R. (Ed.). Methods of soil analysis. 2. ed. Madison: American Society of Agronomy, 1982. p. 643-689. (Agronomy, 9).
LARA CABEZAS, W. A. R.; TRIVELIN, P. C. O.; VICTORIA, R. L.; CAMARGO, P. B.; PICCOLO, M. C. Volatilização de amônia da uréia-15N e aquamônia-15N aplicadas na cultura da cana-de-açúcar em condições de campo. In: CONGRESSO NACIONAL DA SOCIEDADE DOS TÉCNICOS AÇUCAREIROS E ALCOOLEIROS DO BRASIL, 4.; CONVENÇÃO DA ACTALAC, 7., 1987, Olinda, Anais... Olinda: Stab, 1987. p. 5059.
LOBERT, J. M.; SCHSRFFE, D. H.; HAO, W. M.; CRUTZEN, P. J. Importance of biomass burning in the atmospheric budgets of nitrogen-containing gases. Nature, London, v. 346, p. 552554, 1990.
MATTSSON, M.; HAUSLER, R. E.; LEEGOOD, R. C.; SCHJOERRING, J. K. Leaf-atmosphere NH3 exchange in barley mutants with reduced activities of glutamine synthetase. Plant Physiology, Rockville, v. 114, n. 4, p. 13071312, 1997.
MATTSSON, M.; SCHJOERRING, J. K. Characteristics of ammonia emission from barley plants. Plant Physiology and Biochemistry, Paris, v. 34, n. 5, p. 691-695, 1996.
NG KEE KWONG, K. F.; DEVILLE, J. Application of 15N-labeled urea to sugar cane through a drip-irrigation system in Mauritius. Fertilizer Research, Dordrecht, v. 39, p. 223-228, 1994a.
NG KEE KWONG, K. F.; DEVILLE, J. The course of fertilizer nitrogen uptake by rain fed sugarcane in Mauritius. Journal of Agricultural Sciences, Cambridge, Inglaterra, v. 122, p. 385391, 1994b.
OLIVEIRA, M. W. Dinâmica do nitrogênio da uréia (15N) no sistema solo-cana-de-açúcar com ou sem queima da palhada. Piracicaba: USP, 1999. 93 p. Tese de Doutorado.
OLIVEIRA, M. W.; TRIVELIN, P. C. O.; GAVA, G. J. C.; VITTI, A. C. Lixiviação de nitrogênio em solo cultivado com cana-de-açúcar: experimento em lisímetro. STAB Açúcar, Álcool e Subprodutos, Piracicaba, v. 18, n. 2, p. 28-31, 1999.
QUARTA geração de variedades de cana-de-açúcar. Boletim Técnico Copersucar, Piracicaba, dez. 1993. 16 p. Edição Especial.
SCHJOERRING, J. K.; HUSTED, S.; MATTSSON, M. Physiological parameters controlling plant-atmosphere ammonia exchange. Atmospheric Environment, Oxford, v. 32, n. 3, p. 491-498, 1998.
SCHLESINGER, W. H.; HARTLEY, A. E. A global budget for atmospheric NH3. Biogeochemistry, Dordrecht, v. 15, p. 191-211, 1992.
SILVA, L. C. F.; CASAGRANDE, J. C. Nutrição mineral da cana-de-açúcar (macronutrientes). In: ORLANDO FILHO, J. (Coord.). Nutrição e adubação da cana-de-açúcar no Brasil. Piracicaba: IAA/Planalsucar, 1983. p. 77-99.
TRIVELIN, P. C. O. Utilização do nitrogênio pela cana-de-açúcar: três casos estudados com uso do traçador 15N. Piracicaba: USP, 2000. 143 p. Tese de LivreDocência.
TRIVELIN, P. C. O.; LARA CABEZAS, W. A. R.; BOARETO, A. E. Dinâmica do nitrogênio de fertilizantes fluidos no sistema solo-planta. In: VITTI, G. C.; BOARETO, A E. (Ed.). Fertilizantes fluidos. Piracicaba: Potafos, 1994. p. 253268.
TRIVELIN, P. C. O.; RODRIGUES, J. C. S.; VICTORIA, R. L. Utilização por soqueira de cana-de-açúcar de início de safra do nitrogênio da aquamônia-15N e uréia-15N aplicado ao solo em complemento à vinhaça. Pesquisa Agropecuária Brasileira, Brasília, v. 31, n. 2, p. 8999, fev. 1996.
TRIVELIN, P. C. O.; SALATI, E.; MATSUI, E. Preparo de amostras para análise de 15N por espectrometria de massa. Piracicaba: USP-Cena, 1973. 41 p. (Boletim Técnico).
TRIVELIN, P. C. O.; VICTORIA, R. L.; RODRIGUES, J. C. S. Aproveitamento por soqueira de cana-de-açúcar de final de safra do nitrogênio da aquamônia-15N e uréia-15N aplicado ao solo em complemento à vinhaça. Pesquisa Agropecuária Brasileira, Brasília, v. 30, n. 12, p. 1375-1385, dez. 1995.
WEIER, K. L. Sugarcane fields: sources or sinks for greenhouse gas emissions? Australian Journal of Agricultural Research, Collingwood, v. 49, p. 19, 1998.
Paulo Cesar Ocheuze Trivelin(2), Mauro Wagner de Oliveira(3), André Cesar Vitti(4) , Glauber José de Castro Gava(4) e José Albertino Bendassolli(2)
pcotrive[arroba]cena.usp.br
(2) Universidade de São Paulo (USP), Centro de Energia Nuclear na Agricultura (Cena), Caixa Postal 96, CEP 13400970 Piracicaba, SP. Bolsista do CNPq. Email: pcotrive[arroba]cena.usp.br, jab[arroba]cena.usp.br
(3) Universidade Federal de Viçosa, Centro de Pesquisa e Melhoramento de Cana-de-Açúcar, Caixa Postal 342, CEP 35430970 Ponte Nova, MG. E-mail: mwagner[arroba]mail.ufv.br
(4) USP, Cena. Bolsista da Fapesp. Email: acvitti[arroba]cena.usp.br, ggava[arroba]cena.usp.br
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




