 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
O trabalho foi desenvolvido em câmara climatizada (T = 26±0,5oC, UR: 70±10% e fotofase de 14h) utilizando-se lagartas de T. absoluta alimentadas com folhas dos genótipos L. hirsutum f. glabratum (linhagem PI 134417 - resistente) e L. esculentum (cv. Santa Clara - suscetível). As folhas foram tratadas com suspensão do fungo entomopatogênico Beauveria bassiana (Bb).
Folhas dos dois genótipos foram coletadas de plantas com 15 a 20 cm de altura, mantidas em casa de vegetação e colocadas em placas plásticas (6,5 x 2,5 cm) e infestadas com lagartas recém-eclodidas. Visando à obtenção de insetos em diferentes estágios de desenvolvimento, as lagartas foram mantidas alimentando-se de folhas da cv. Santa Clara e PI 134417, pelos seguintes períodos de tempo, respectivamente: a) um dia para o 1o ínstar; b) quatro e cinco dias para atingir o 2o ínstar; c) seis e oito dias para atingir o 3o ínstar, e d) 10 e 12 dias para atingir o 4o ínstar (Giustolin 1996). Ao atingirem as referidas idades, as lagartas foram transferidas para folhas tratadas com suspensão de conídios de Bb. Foi ainda realizado um experimento em que as lagartas não receberam qualquer alimento antes de receberem folhas tratadas. Ao todo foram instalados cinco experimentos, sendo quatro com lagartas previamente alimentadas por diferentes períodos e um com lagartas recém-eclodidas.
O isolado de B. bassiana foi cedido pelo laboratório de Patologia de Insetos do Departamento de Entomologia, Fitopatologia e Zoologia Agrícola (ESALQ/USP). Foi isolado de Solenopsis invicta Buren (Cuiabá, MT) e armazenado na micoteca do referido laboratório sob o número 447. A suspensão foi preparada com conídios, 100% viáveis, mais espalhante adesivo (Tween 20® a 1%), utilizando-se a concentração de 1,0x106 conídios/ml, correspondente a CL70 para lagartas recém-eclodidas (Giustolin 1996). Para o tratamento com Bb, discos foliares (diâmetro de 2,1 cm) dos dois genótipos foram imersos na suspensão de conídios, por cinco segundos, em câmara de fluxo laminar, onde permaneceram até a evaporação do excesso de líquido. Os discos foram transferidos para placas plásticas (6,5 x 2,5 cm) forradas com papel-filtro esterilizado e umedecido e, então, infestados com lagartas nos diferentes estágios de desenvolvimento. Discos dos respectivos genótipos imersos em água foram utilizados como testemunha.
Para cada estágio de desenvolvimento, cada um deles constituindo um experimento, foram utilizados os seguintes tratamentos: Santa Clara + Bb; Santa Clara + água; PI 134417 + Bb e PI 134417 + água. O tratamento com Bb foi feito apenas uma vez no início do experimento, sendo, sempre que necessário, fornecidas novas folhas (não tratadas) às lagartas. Para cada um dos cinco experimentos, utilizou-se o delineamento inteiramente casualizado com quatro tratamentos e 20 repetições (com três lagartas cada).
As placas foram observadas diariamente, avaliando-se a mortalidade larval. Os dados de cada um dos experimentos, que correspondeu a cada uma das idades das lagartas, foram submetidos a análise de variância, sendo as médias comparadas pelo teste de Tukey (P < 0,05), adotando-se esquema fatorial, considerando-se os fatores genótipo e patógeno.
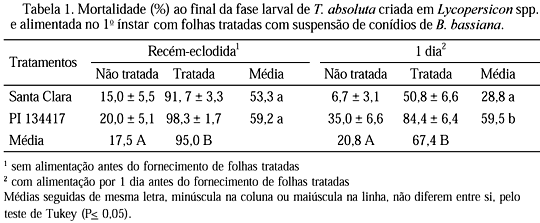
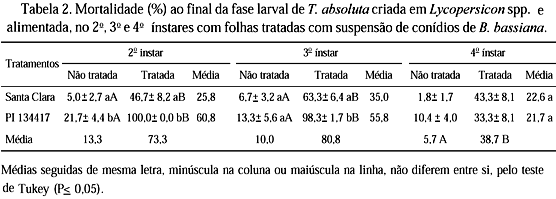
Nos experimentos com lagartas de T. absoluta recém-eclodidas (sem alimentação antes do fornecimento de folhas tratadas) e de 4o ínstar, a mortalidade larval deveu-se apenas ao fator patógeno (Bb), constatando-se maior mortalidade nas parcelas tratadas independente dos genótipos (Tabelas 1 e 2). Nesses dois experimentos não foi observada a ocorrência de sinergismo entre os fatores genótipo e patógeno. Quando as lagartas foram alimentadas com folhas tratadas logo após a eclosão, cerca de 25% de mortalidade larval foi constatada já no dia seguinte. Até o 4o dia de avaliação, a mortalidade provocada pelo patógeno foi maior nas lagartas criadas em 'Santa Clara' que na PI 134417. A partir do 5o dia, os valores de mortalidade (cerca de 80%) foram semelhantes nos dois genótipos (Fig. 1A). Quando as lagartas receberam folhas tratadas no 4o ínstar, a eficiência do patógeno foi semelhante nos dois genótipos, ocorrendo aumento gradativo na mortalidade a partir do 4o dia, não chegando, no entanto, a atingir 45%, ao final da fase larval (Fig. 1E). Nesses dois experimentos, quando as lagartas receberam folhas sem Bb, a mortalidade larval, nos dois genótipos, foi relativamente baixa ao longo do período de avaliação (Figs. 1A e 1E).
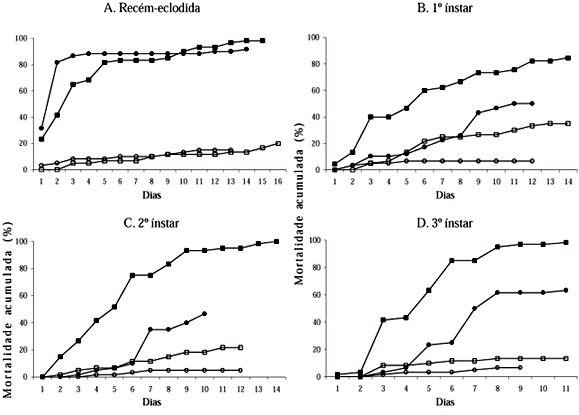
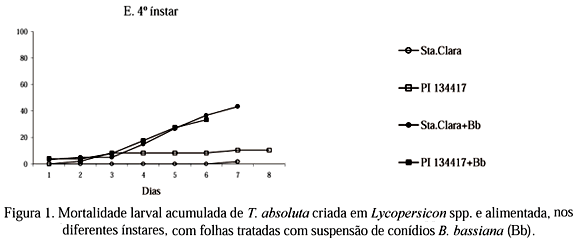
A alta mortalidade larval no experimento com lagartas recém-eclodidas alimentadas nos genótipos tratados contrastou com os resultados observados nesses mesmos tratamentos no experimento com lagartas de 4o ínstar. Isso pode ter ocorrido em razão da maior suscetibilidade das lagartas mais jovens e ou devido ao comportamento desse inseto minador que, logo após a eclosão, raspa a folha durante 20 a 45 min. antes de penetrar no mesofilo (Coelho & França 1987), possibilitando um contato mais prolongado com os conídios de Bb. Em relação às lagartas que receberam folhas tratadas apenas no 4o ínstar, a baixa mortalidade ao final da fase pode ter ocorrido devido a concentração insuficiente de conídios de Bb, considerando-se o maior tamanho do inseto e ou ao fato de o inseto, no último ínstar, ter liberado a exúvia antes da penetração do tubo germinativo do patógeno. Esta última hipótese é respaldada pelos estudos de Fargues (1972), que mencionou que a liberação da exúvia é um obstáculo natural para a infecção e a contaminação do inseto por Bb.
No experimento em que as lagartas de 1o ínstar foram alimentadas nos genótipos durante um dia, antes de receberem folhas tratadas, a mortalidade larval deveu-se aos fatores genótipo e patógeno. Assim, aquelas criadas na PI 134417 apresentaram maior mortalidade ao final da fase larval que as criadas na cv. Santa Clara, independente da presença do entomopatógeno. Por outro lado, as lagartas alimentadas com folhas tratadas com Bb apresentaram maior mortalidade que as alimentadas com folhas não tratadas, independente dos genótipos (Tabela 1). Nesse caso, não foi observada interação entre os fatores de resistência com Bb, ocorrendo, portanto, efeitos independentes e aditivos. Os dados de mortalidade acumulada nesse experimento (Fig. 1 B) evidenciam que cerca de 50% das lagartas morreram cinco dias após iniciarem a alimentação na PI 134417 tratada com Bb, enquanto que em 'Santa Clara' esse mesmo percentual ocorreu 11 dias após, indicando maior virulência de Bb na PI 134417. Nos genótipos não tratados, a mortalidade larval foi mais baixa em relação às respectivas parcelas tratadas, principalmente em 'Santa Clara' onde, ao final da fase larval, a mortalidade atingiu apenas 7%, contra 35% na PI 134417.
Nos experimentos em que as lagartas receberam folhas tratadas no 2o e no 3o ínstar, foi constatada interação significativa dos fatores genótipo e patógeno. A mortalidade larval na PI 134417 foi maior que na cv. Santa Clara, tanto com folhas tratadas com Bb (para os dois ínstares), como com folhas não tratadas (lagartas de 2o ínstar) (Tabela 2). O efeito do patógeno foi bastante evidente nos dois ínstares, com maior mortalidade larval nas folhas tratadas nos dois genótipos (Tabela 2). No tratamento PI 134417 mais Bb, a mortalidade foi maior que o somatório das mortalidades causadas pelos fatores genótipo e Bb isoladamente, ocorrendo, nesse caso, sinergismo dos dois fatores. Com lagartas que receberam folhas tratadas no 2o ínstar (Fig. 1C), a mortalidade acumulada na PI 134417, no 5o dia após a aplicação do patógeno, já era de 50%, atingindo 100% ao final do experimento. Na cv. Santa Clara, entretanto, a mortalidade não atingiu 50%, mesmo ao final do experimento (10 dias após a aplicação de Bb). Para lagartas cuja alimentação em folhas tratadas ocorreu no 3o ínstar (Fig. 1D), cerca de 50% de mortalidade nas parcelas tratadas foi observada após quatro dias na PI 134417 e após sete dias em 'Santa Clara'. Nesses dois experimentos (lagartas de 2o e 3o ínstar), a mortalidade larval nos genótipos não tratados foi relativamente baixa ao longo de todo o período de avaliação (Figs. 1C e 1D).
Nos experimentos com as lagartas alimentadas com folhas tratadas nos 1o, 2o e 3o ínstares, as altas mortalidades ocorridas no tratamento PI 134417 mais Bb em relação à 'Santa Clara' mais Bb (Tabelas 1 e 2), provavelmente, se deveram aos fatores de resistência da PI 134417, que teriam estressado os insetos, tornando-os mais suscetíveis à doença. Hare & Andreadis (1983) também observaram maior suscetibilidade de L. decemlineata a Bb nos genótipos resistentes de Solanum spp. e L. esculentum em relação aos genótipos suscetíveis. Já, com lagartas alimentadas com folhas tratadas desde a eclosão, essa diferença entre genótipos não ficou caracterizada, pois nos dois genótipos as mortalidades foram bastante elevadas, atingindo índices acima de 90% (Tabela 1).
Em nenhum dos experimentos, foi observada variação na suscetibilidade do inseto em função dos ínstares em que o mesmo foi submetido ao patógeno. Fargues (1972) observou que larvas de L. decemlineata em ínstares mais avançados são menos suscetíveis ao fungo e que nos primeiros ínstares elas suprimem ou superam a infecção através da liberação da exúvia antes que as hifas alcancem o hemocele. O mesmo autor observou, ainda, maior mortalidade das larvas tratadas com Bb, três dias antes de sofrerem ecdise. Li et al. (1995) também observaram diferença na suscetibilidade de Choristoneura rosaceana (Harris) em diferentes ínstares a Bacillus thuringiensis, sendo mais suscetível durante o 4º ínstar e menos no 6º ínstar.
Nos tratamentos sem aplicação de patógeno, observou-se que a linhagem PI 134417 causou maior mortalidade larval que a cv. Santa Clara. A mortalidade causada pela linhagem foi maior nos primeiros ínstares, diminuindo com o desenvolvimento, demonstrando que o inseto é mais suscetível a esse genótipo no início da fase larval (Tabelas 1 e 2). Estes dados confirmam novamente o efeito prejudicial da linhagem PI 134417 sobre as lagartas de T. absoluta e, ao mesmo tempo, indicam que o referido patógeno é promissor no controle do inseto. Entretanto, efeitos adversos dos fatores químicos dos genótipos sobre Bb, não foram observados em nenhum dos experimentos.
Teresinha A. Giustolin, José D. Vendramim, Sérgio B. Alves E Solange A. Vieira
sebalves[arroba]esalq.usp.br
Depto. de Entomologia, Fitopatologia e Zoologia Agrícola, ESALQ/USP
Caixa postal 9, 13418-900, Piracicaba, SP
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




