 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
O estudo foi realizado no Instituto de Zootecnia em Nova Odessa, SP. Foram utilizados 48 leitões, machos castrados e fêmeas da mesma procedência genética, com peso inicial de 5,5 ± 0,23 kg. Os animais, provenientes de cruzamentos entre reprodutores e matrizes comerciais, foram distribuídos em delineamento experimental de blocos ao acaso, com quatro tratamentos e seis repetições. Na formação dos blocos, foram adotados os pesos iniciais.
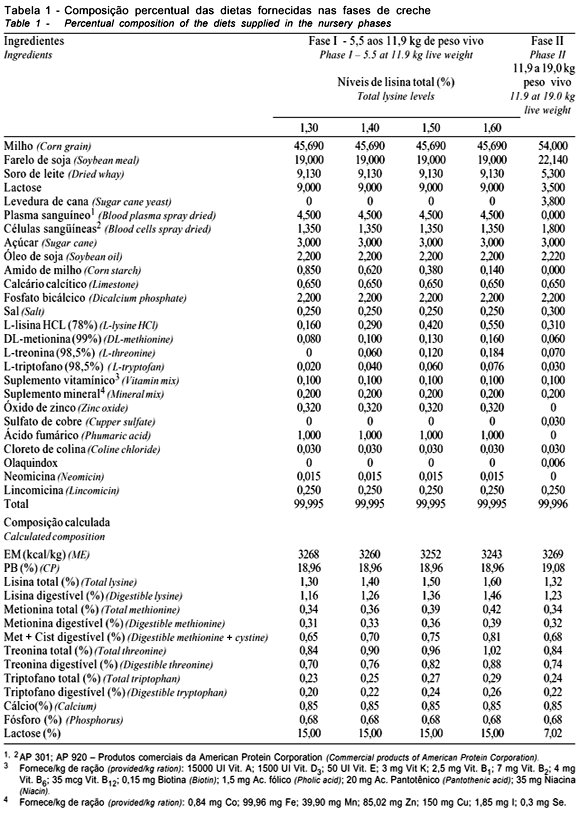
Os tratamentos (Tabela 1) corresponderam a dietas com níveis 1,30; 1,40; 1,50 e 1,60% de lisina total, oferecidas durante 17 dias (fase inicial-1). Posteriormente, receberam a mesma dieta durante 13 dias (fase inicial-2) até o final do período de creche. As relações estabelecidas entre os principais aminoácidos ocorreram conforme recomendações do NRC (1998). A alimentação era à vontade e o controle do desenvolvimento ocorreu no final de cada fase.
Um grupo adicional de seis leitões foi abatido no dia do desmame. Os animais (três machos castrados e três fêmeas) encontravam-se na mesma faixa de peso dos demais. Os dados médios obtidos nesta etapa serviram como valores comparativos da composição química corporal, sendo utilizados na determinação das taxas de deposição protéica e lipídica ao final do experimento.
Ao final de cada fase, após 16 horas de jejum, 24 leitões (seis/tratamento) foram abatidos para determinação da composição química nas frações corporais: sangue, vísceras e carcaça; no corpo vazio, bem como taxas diárias de deposição protéica e lipídica. O abate ocorreu pelo método de sangria após insensibilização elétrica.
Definiu-se como vísceras: trato digestivo e urinário vazios, glândulas anexas, órgãos reprodutores, coração, fígado, baço, pulmão, rins e gordura perirenal. Considerou-se como carcaça o suíno sangrado e eviscerado. A carcaça incluía cabeça, pés, unhas e pêlos. O corpo vazio foi definido como diferença entre o peso vivo em jejum e os conteúdos estomacal, intestinal e da bexiga (Quiniou & Noblet, 1995; Trindade Neto et al., 2001).
O sangue foi coletado, pesado e retiravam-se três amostras de 80 mL por animal. As vísceras foram retiradas, esvaziadas e pesadas. As vísceras e a carcaça foram embaladas e mantidas a -10ºC até o processamento, quando foram reduzidas ao estado pastoso por intermédio de um moedor de carne (Herman P-33A-3-789), retirando-se quatro amostras em placas de Petri.
As amostras de sangue, de vísceras e de carcaça foram liofilizadas pelo método de sublimação, no Instituto de Tecnologia de Alimentos - Campinas, SP. O liofilizador foi o Stokes, sistema de vácuo a 3 mmHg de pressão máxima em temperaturas inicial de -15ºC e final de 10ºC. As amostras liofilizadas foram moídas com gelo seco para subseqüentes análises bromatológicas.
As análises bromatológicas foram feitas em duplicatas. Em cada fração corporal, determinaram-se os teores de matéria seca, proteína bruta, extrato etéreo e cinzas. A matéria seca considerada foi a liofilizada, contudo, para correção do extrato etéreo, da proteína bruta e da matéria mineral, também foi utilizada a obtida em estufa a 105ºC, uma vez que, durante a manipulação, poderia haver alterações da umidade inicial. Os dados de vísceras, sangue, carcaça e no corpo vazio foram expressos na matéria natural, liofilizada e corrigida pela matéria seca. As taxas diárias de deposição avaliadas foram: proteína, lipídeo e água na carcaça e no corpo vazio.
As características determinadas na composição química corporal foram submetidas à análise de regressão por polinômios ortogonais, por intermédio do SAS (SAS, 1996).
Os dados das frações corporais e do corpo vazio na fase inicial-1 constam na Tabela 2.
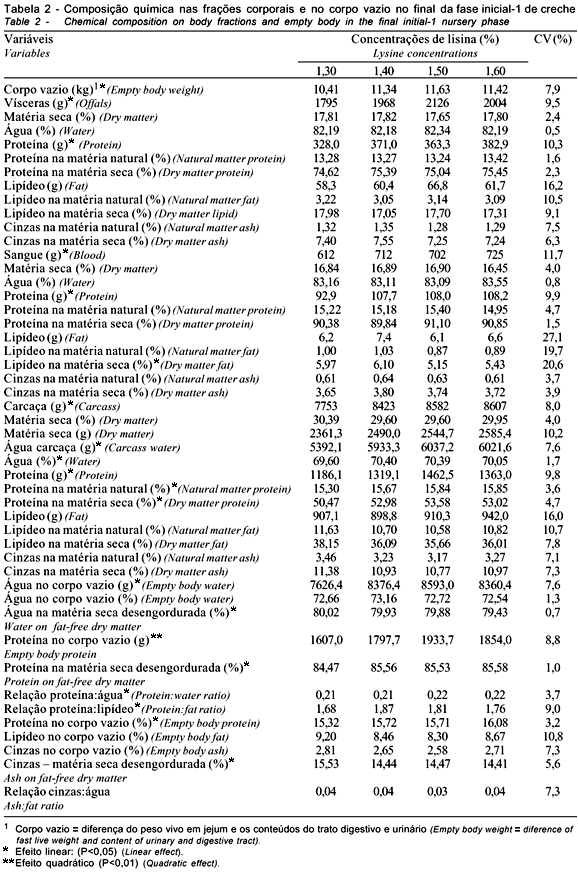
Aumento da concentração dietética de lisina propiciou resposta linear (P<0,05) ascendente no corpo vazio. O mesmo efeito foi observado no peso das frações corporais: vísceras (P<0,03); sangue (P<0,03); e carcaça (P<0,04). A resposta linear nessas variáveis correlacionou-se com o aumento do peso corporal, em função do crescente e linear (P<0,08) ganho de peso, em resposta à elevação da concentração de lisina na dieta (Tabela 3).
Separadamente, ao quantificar os componentes químicos das frações corporais e do corpo vazio, foram observadas respostas diferentes para os níveis nutricionais estudados.
Houve resposta linear na quantidade (g) de proteína das vísceras (P<0,08) e do sangue (P<0,02), em decorrência do aumento dos níveis de lisina na dieta, que pode ter correspondido à variação do peso dos animais e respectivas frações, e do tamanho de órgãos como fígado e rins. O aumento desses órgãos decorreria também da maior atividade metabólica, em razão do aumento do fluxo de nutrientes sangüíneos proveniente do crescente aporte de lisina e dos demais aminoácidos dietéticos, segundo a estimativa de consumo apresentada na Tabela 3. O fígado e os rins possuem papel importante na regulação dos níveis de aminoácidos no plasma sangüíneo e na síntese de glicose, proveniente do excesso de aminoácidos. A proteína (aminoácidos) promove o crescimento desses órgãos mais que o resto do corpo, sugerindo-se hipertrofia e hiperplasia (Chan & Hargrove, 1993).
Na carcaça, a elevação do nível de lisina na dieta propiciou resposta linear (P<0,02) ascendente para a quantidade (g) de água e resposta quadrática (P<0,04) para a de proteína, que aumentou até o nível estimado de 1,51%.
Aumento da quantidade de água está relacionado à maior síntese protéica e essa, por sua vez, ao maior ritmo no aumento de peso (Bondi, 1988). Durante o crescimento, a síntese protéica destina-se, preferencialmente, à formação muscular em uma relação direta com o teor de água. Na carcaça, a água corresponde a ¾ do tecido magro e a proteína, apenas ¼ (Fuller & Wang, 1990; Claus & Weiler, 1994).
Em relação à quantidade de proteína na carcaça, o nível ótimo de lisina seria 1,51%, entretanto, essa variável não deve ser avaliada isoladamente na determinação do melhor nível de lisina. Há relações da síntese protéica com a deposição de água e de lipídios, que pode variar no conjunto das frações corporais ou corpo vazio. Essas variações ocorrem na dependência da relação entre os nutrientes dietéticos, sobretudo nas fases de maior crescimento do suíno, caracterizado pelo acúmulo de massa protéica corporal, como ocorre após o desmame. Ao considerarem a máxima deposição de proteína, Dunshea et al. (2000) sugeriram 1,50% de lisina total para leitões entre 5 kg e 10 kg de peso vivo, com base na relação ideal entre aminoácidos e no conceito da proteína ideal.
Transformados em valores relativos percentuais, o aumento dos níveis de lisina nas dietas implicou em concentração linear ascendente de proteína na carcaça, expressos na matéria natural (P<0,09) e na matéria seca (P<0,08). Uma vez que a proteína é o principal componente da matéria seca de órgãos e tecidos (Bondi, 1988), o aumento da concentração dietética de lisina e dos demais aminoácidos teria permitido maior eficiência metabólica, caracterizada na pequena variação da composição química da carcaça. Esse efeito está de acordo com as observações de Henry et al. (1992) e Susenbeth (1995), quando verificaram que o aumento da concentração de lisina beneficia o desenvolvimento das frações ou compartimentos protéicos do corpo.
Segundo Friesen et al. (1996), a síntese protéica não estaria associada integralmente à deposição muscular durante o crescimento, mas o destino preferencial seria a deposição muscular esquelética. Em estudo com leitões até a quarta semana de idade, Davis et al. (1996) concluíram que a intensidade da síntese protéica muscular esquelética é um reflexo da alimentação. No presente estudo, o aumento do teor de proteína na carcaça sugere níveis de lisina não inferior a 1,50%.
Na soma das frações corporais aferidas como corpo vazio, verificou-se efeito quadrático para a quantidade (g) de proteína (P<0,05) e de água (P<0,07). O nível ótimo de lisina total para a quantidade de proteína e de água seria de 1,52 e 1,50%, respectivamente. No segundo caso, embora em pequena quantidade, o tecido adiposo contém água (Bondi, 1988) e contribui na totalização dos resultados das frações corporais. Assim, a proteína seria indicativo mais preciso, como observado nos resultados encontrados na matéria seca desengordurada (Tabela 2).
Confirmando as respostas quantitativas no corpo vazio, os valores relativos à porcentagem de proteína apresentaram aumento, em função dos níveis dietéticos de lisina; não obstante, as respostas foram lineares ascendentes na matéria natural (P<0,02) e na matéria seca desengordurada (P<0,04). A variação na composição protéica de 15,32% a 16,08% na matéria natural e de 84,47% a 85,58% na matéria seca desengordurada é indicativo de que o aumento da lisina e dos demais aminoácidos na dieta propiciaria maior eficiência metabólica, em relação ao acréscimo de proteína corporal. Essa variação está de acordo com as informações de Whittemore (1993), de que o conteúdo de proteína corporal é mais estável em relação ao conteúdo de gordura, variando entre os extremos de 14 e 18%, assumindo-se, na média, 16%. Segundo esse autor, o conteúdo de carne magra do corpo varia inversamente ao tecido gorduroso, decorrente, em parte, da variação do conteúdo de água na carne, que estaria relacionada à redução do anabolismo protéico, concomitantemente ao aumento da deposição de gordura, com o avanço da idade e do peso do suíno, razão pela qual a eficiência do crescimento está associada ao acúmulo de massa muscular. Normalmente, o músculo possui 70% a 75% de água, 5% a 15% de lipídeo e 20% a 25% de proteína. Em suínos muito jovens, quando o processo de síntese protéica muscular é mais intenso, o conteúdo de água pode atingir concentrações acima de 80%, enquanto, no adulto, pode estar bem abaixo de 70%.
Quando foi considerada a matéria seca desengordurada, o percentual de água no corpo vazio tendeu a redução (P<0,08) linear. Inferindo sobre a composição corporal do suíno, Whittemore (1993) informou que o tecido gorduroso possui entre 10 e 25% de água, 2% de proteína e entre 70 e 80% de lipídeo e, durante o crescimento do animal, a proporção de gordura é muito variável, porém essa variação depende dos níveis de nutrientes supridos, além dos fatores sexo e genótipo.
O teor de cinzas na matéria desengordurada diminuiu (P<0,04) de forma linear, em resposta à crescente concentração da lisina na dieta. Não há justificativa técnica para a variação observada, embora tenha havido aumento do teor protéico corporal. Segundo Whittemore (1993), a relação entre cinzas e proteína é relativamente constante e as cinzas correspondem na média 3% do peso vivo ou 20% da proteína, pois os ossos são usados como estrutura suporte para a musculatura. No segundo caso, essa relação é confirmada na fração carcaça.
Aumento do teor protéico na matéria seca desengordurada ficou melhor caracterizado quando se estabeleceu a relação entre proteína e água, cujo aumento (P<0,03) ratifica a maior eficiência na síntese e deposição de proteína (anabolismo), em resposta ao aumento da lisina. A resposta obtida na matéria seca desengordurada indica que a composição química corporal pode variar em pequenas quantidades. Essa pequena variação poderia decorrer do aporte combinado dos limitantes e demais aminoácidos, no metabolismo protéico do suíno.
Da mesma forma, a relação entre proteína e lipídeo é um indicativo da eficiência da síntese protéica, em resposta ao nível de lisina na dieta. Nesse caso, o efeito quadrático (P<0,09) indica como ponto ótimo o nível de 1,46% de lisina total. Ainda que seja indicativo e as variações sejam pequenas, isoladamente, essa variável não pode ser considerada na determinação do melhor nível de lisina. Os resultados da literatura são divergentes com relação aos efeitos da lisina na composição química corporal do suíno livre de gordura.
Segundo Susenbeth & Keitel (1988) e Trindade Neto et al. (2001), os dados de composição química corporal livre de gordura variam pouco. Entretanto, Whittemore (1993) informa que a variação da gordura corporal depende dos níveis de nutrientes supridos, enquanto Lawrence & Fowler (1997) sugerem o acúmulo de proteína celular, portanto, poderia haver pequena variação.
A maior quantidade e concentração de proteína nas frações corporais e no corpo vazio indica o aumento da eficiência na utilização da lisina para a síntese protéica nos leitões, em resposta à elevação dos níveis dos aminoácidos. Os resultados ratificam observações de Noblet et al. (1987), Henry et al. (1992), Susenbeth (1995) e Trindade Neto et al. (2001) de que o aumento dos níveis de lisina estimula o desenvolvimento das frações ou compartimentos protéicos do corpo. Essa situação ocorre quando o suíno tem potencial genético para alta deposição de massa muscular e a demanda para atender essa máxima eficiência de utilização não está sendo suprida, considerando que o animal esteja em condições desejáveis de saúde (Williams et al., 1997).
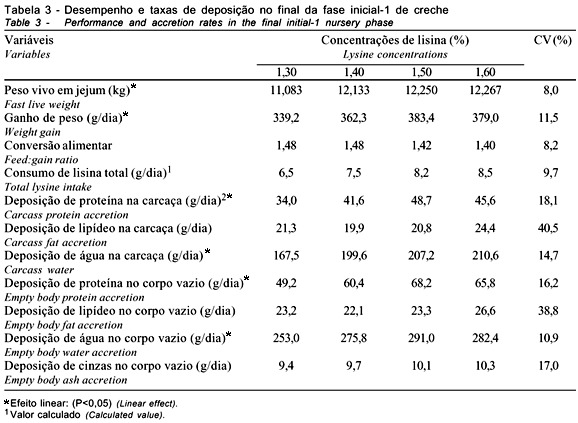
Os resultados de desempenho e decomposição do ganho de peso dos leitões na fase inicial-1 são apresentados na Tabela 3.
Ao elevar a concentração de lisina na dieta, observou-se o aumento linear (P<0,08) no ganho de peso. As médias de ganho, ajustadas pela equação de regressão, seriam: 345,0; 359,0; 373,0 e 387,0 g/dia, respectivamente, para os níveis de lisina estudados. Essa estimativa sugere a maior eficiência na utilização da dieta, caracterizada pela melhora linear (P<0,03) da conversão alimentar. Considerando o desempenho ponderal dos leitões, os efeitos lineares observados mostram que os leitões respondem favoravelmente até o nível 1,60% de lisina total na dieta, uma vez mantidas as relações entre os principais aminoácidos. Neste estudo, procurou-se manter as relações entre lisina, metionina, treonina e triptofano nas dietas experimentais, conforme recomendação do NRC (1998).
A ingestão calculada de lisina teria o mesmo comportamento ascendente e no nível 1,60% o consumo seria 4,93 g do aminoácido por megacaloria de energia metabolizável. O valor estimado estaria próximo à recomendação do NRC (1998) para consumo diário de 500 g de ração, mas as necessidades para maximizar o desempenho não estariam, precisamente, definidas. Van Der Peet-Schwering et al. (1999), em uma revisão, observaram que a máxima deposição de proteína somente ocorre quando a ingestão de energia e dos aminoácidos não são limitadas. No caso de limitação dos aminoácidos, a deposição de proteína será reduzida. Com base nos resultados desta pesquisa, sugere-se que sejam desenvolvidos estudos com níveis acima e abaixo do máximo proposto, para determinar o ponto ótimo de lisina, dentro do conceito de proteína ideal, uma vez conhecidas as demandas de energia para o material genético a ser utilizado.
Ao decompor o ganho de peso, observou-se efeito quadrático (P<0,06) dos níveis de lisina sobre as taxas diárias de deposição protéica na carcaça e efeito linear ascendente (P<0,01) no corpo vazio, com tendência à resposta quadrática (P<0,10) (Figura 1), não havendo efeitos na deposição lipídica.
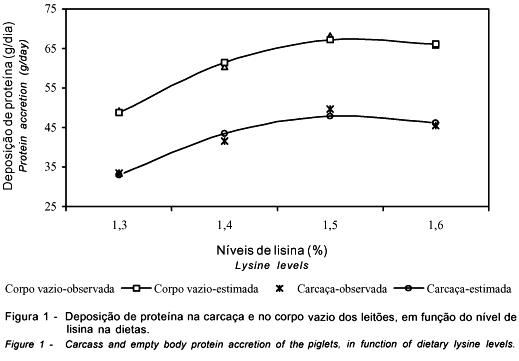
Aumento da deposição de proteína, em resposta à concentração de lisina e demais aminoácidos, evidencia gradativa eficácia na utilização dos nutrientes dietéticos para o metabolismo no processo de síntese protéica e acúmulo de massa muscular esquelética, uma vez que foi mínima a variação entre os níveis de energia nas dietas experimentais. Os valores médios de deposição protéica na carcaça e no corpo vazio foram superiores aos determinados por Trindade Neto et al. (2001), com animais abatidos aos 11,6 kg de peso vivo, quando variaram as concentrações de lisina de 0,85% a 1,25% em dietas isonitrogenadas, ausentes de produtos lácteos e da relação ideal entre os principais aminoácidos. Neste estudo, além da proteína ideal, as dietas continham produtos lácteos, plasma e hemácias liofilizados, e níveis de lisina mais elevados, favorecendo o consumo de ração. No estudo anterior, os autores observaram que a característica da dieta afetou o consumo, aumentando o tempo para que os animais atingissem o peso esperado.
Ainda que não fossem detectados efeitos significativos, em decorrência dos elevados coeficientes de variação (40,5 e 38,8%, respectivamente), a deposição lipídica na carcaça e no corpo vazio teve comportamento semelhante, porém em quantidades inferiores à proteína. Susenbeth & Keitel (1988) descreveram grandes variações na taxa de deposição lipídica. Essa variação na deposição decorre da ação conjugada de enzimas, como a lípase lipoprotéica e triacilglicerol, no processo de biosíntese (lipogênese), que se intensifica após o desmame e com o avanço da idade. Juntamente com a deposição lipídica, há aumento de colesterol, de ácidos biliares e de fosfolipídeos, como resposta à alimentação, mas, segundo Friesen et al. (1995), o controle homeostático no metabolismo do colesterol não seria afetado pela lisina dietética.
Houve aumento linear na deposição de água na carcaça (P<0,02) e água total no corpo vazio (P<0,07). Aumento de proteína e de água na carcaça corroborou as observações de Friesen et al. (1996) de que, no processo do crescimento, a lisina é direcionada, principalmente, para síntese e acúmulo de massa muscular. A variação do consumo estimado de lisina (6,5 a 8,5 g) foi fundamental na quantidade diária de deposição protéica, conforme também verificaram Schinckel & Lange (1996) e Trindade Neto (1999), quando trabalharam com suínos em plena fase de crescimento. Como ocorreu em outras variáveis, as estimativas de consumo do presente estudo aproximam-se da recomendação de 6,75 g/dia do aminoácido, segundo o NRC (1998). Ratifica-se, porém, a necessidade da aplicação de maior número de níveis, para provável e mais acurada determinação do valor de lisina que maximize o desempenho, visto que recursos adicionais da estatística não puderam ser eficientemente aplicados na avaliação dos dados experimentais.

O progressivo acúmulo protéico confirmou os resultados obtidos por Williams et al. (1997) e Trindade Neto et al. (2001), quando trabalharam com categorias de suínos semelhantes. Os resultados da fase inicial-1 corroboram aqueles observados na literatura, em que a ingestão diária de lisina determina a intensidade da síntese protéica, até certa concentração, porém insuficiente aporte energético para realização desse processo limitaria a utilização do aminoácido. Nesse caso, parte do pool de aminoácidos, cuja referência é a lisina, seria utilizada para produção de energia. Embora com base nas recomendações do NRC (1998), os níveis de energia empregados na elaboração das dietas experimentais, provavelmente, não atenderam às demandas de manutenção e crescimento dos leitões. Segundo Susenbeth (1995) e Van Lunen & Cole (1998), a relação de dependência entre ingestão de energia e utilização da lisina caracteriza-se quando a energia limita a retenção de proteína. Na quantificação dos efeitos do consumo de energia sobre a retenção protéica, a maior dificuldade seria o potencial de crescimento do animal e seu efeito no uso da energia. O potencial de crescimento é definido como retenção de proteína, quando a oferta de energia ocorre sem qualquer limitação dos demais nutrientes, em condições desejáveis de ambiente.
Os dados da fase inicial-2 são apresentados nas Tabelas 5 e 6. Não foram observados efeitos residuais das concentrações de lisina ao final da fase inicial-2. Os animais que, anteriormente, tiveram menor desempenho recuperaram-se quando receberam a dieta-padrão. Em relação à concentração dos componentes químicos das frações corporais e do corpo vazio, bem como as taxas diárias de deposição, também não foram observadas diferenças residuais dos tratamentos aplicados na fase inicial-1.
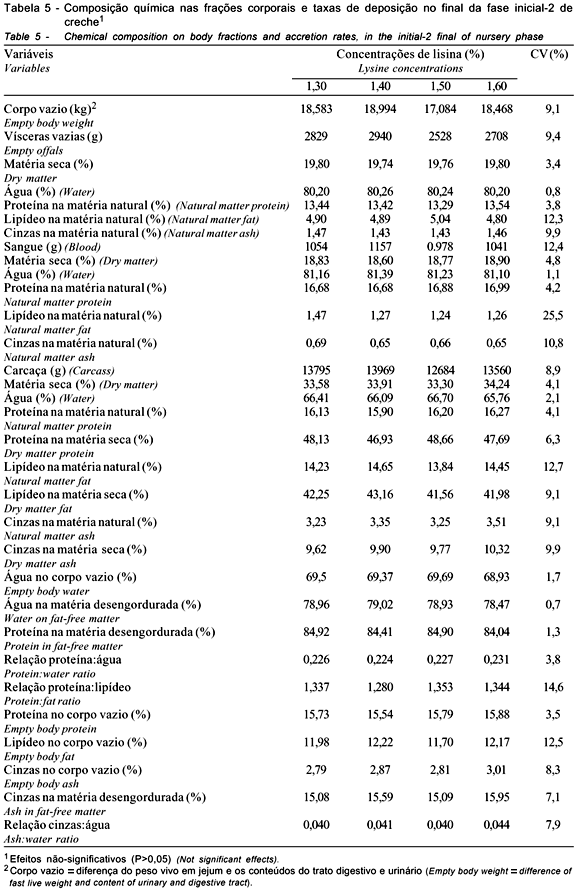
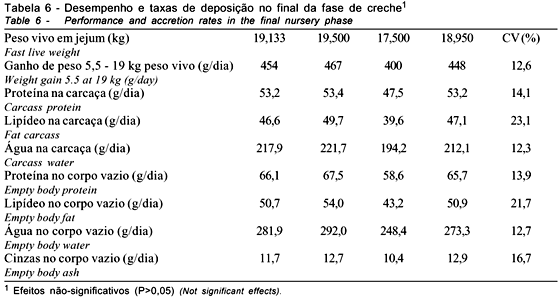
Os animais que se encontravam com menor peso na fase anterior foram mais eficientes na utilização da dieta oferecida na fase subseqüente aos tratamentos experimentais. A recuperação da deposição protéica observada correspondeu à variação negativa, não-significativa, do ganho de peso daqueles animais que anteriormente tiveram melhor desempenho. A inversão de respostas pode caracterizar provável restrição nutricional aos leitões que receberam menores níveis de lisina na fase inicial-1. Essa restrição anterior teria limitado o aporte de aminoácidos para o acúmulo de massa muscular, como foi observado nas variáveis químicas e, principalmente, no ganho de peso e nas taxas diárias de deposição protéicas (Tabelas 2 e 3).
Os resultados obtidos com o genótipo testado diferem daqueles de Van Lunen & Cole (1998), que verificaram aumento do ganho protéico (1,14 a 2,00%), fornecendo concentrações de lisina para leitões entre 9 e 25 kg de peso vivo, e confirmaram a limitação da síntese protéica para a ingestão diária de lisina.
Considerando a diversidade de dados encontrados para diferentes genótipos, acredita-se que o verdadeiro potencial de desempenho possa estar além do limite máximo do apetite. Provavelmente, esse é o fator mais restritivo da deposição muscular nos suínos melhorados para produção de carne magra (Close, 1994). Conforme a pequena variação da porcentagem de proteína na matéria natural, da deposição protéica e deposição de água na carcaça e no corpo vazio, durante o fornecimento dos tratamentos, verifica-se aumento no processo anabólico protéico, carreada para o incremento da massa muscular.
Acerca da recuperação dos leitões no período subseqüente aos tratamentos experimentais, entende-se que há um limite fisiológico, não estabelecido pela pesquisa, no qual o déficit de nutrientes dietéticos, por determinado período, não compromete o desempenho seguinte do animal, quando o suíno pode recuperar o limitado desempenho de fase curta (imediatamente anterior), se as demais condições de meio forem satisfatórias.
Outra hipótese seria as concentrações nutricionais para a segunda fase excederem às reais necessidades dos leitões. Nesse caso, aqueles em déficit seriam mais eficazes na utilização dos nutrientes dietéticos e os que haviam tido maior ganho catabolisariam parte desses nutrientes. Na segunda situação, o catabolismo dos nutrientes excedentes teria implicações negativas no desempenho.
O ideal seria trabalhar, no mínimo, com cinco níveis de lisina, para permitir melhor exploração estatística dos dados e inferências mais precisas sobre o efeito do nível de lisina.
As respostas favoráveis do aumento da concentração de lisina até o nível de 1,6% na dieta de leitões entre 5,5 e 11,9 kg recomendam a realização de novos estudos utilizando níveis de lisina superiores aos empregados neste trabalho, combinados com maiores níveis de energia metabolizável, a fim de estabelecer a eficiência máxima de deposição protéica.
Messias Alves da Trindade NetoI; Izabel Marin PetelincarII; Dirlei Antônio BertoIII; Eliana Aparecida SchammassII; Kátia Sardinha BisinotoIV; Fabiana Ribeiro CaldaraIV
messiasn[arroba]usp.br
IProfessor Doutor do Departamento de Nutrição e Produção Animal da FMVZ-USP, Campus de Pirassununga, Rua Duque de Caxias Norte, 225, CEP:13630-000 Pirassununga SP (
IIInstituto de Zootecnia Rua Heitor Penteado 56, Nova Odessa, SP, CEP: 13460-000
IIIProfessor Doutor do Departamento de Produção e Exploração Animal, FMVZ, UNESP/Botucatu, SP
IVAluno de pós-graduação - Departamento de Zootecnia da UNESP/Botucatu, FCA
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




