 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
Os dados para este trabalho foram obtidos dos livros de escrituração zootécnica do NPZGJRA/IZ/APTA/SAA-SP, desde a formação do rebanho Mantiqueira, na década de 50. Foram incluídos somente registros de vacas com um ou mais partos e de bezerras e novilhas para futura reposição no rebanho, assim como de todos os machos selecionados como reprodutores, totalizando 2.070 animais.
O núcleo básico foi formado pela aquisição de vacas e touros de quatro rebanhos de Minas Gerais e de um rebanho de São Paulo e, desde então, o rebanho foi fechado, não sendo mais permitida a introdução de material genético de qualquer origem. Da formação do rebanho até o ano de 1974, simplesmente procurou-se ampliar o número de animais, sem que houvesse um programa de acasalamento entre animais não aparentados. Apenas em 1974, elaborou-se tal programa, utilizando-se quatro touros de origens distintas ("linhagens") e tendo como critério a não existência de avós em comum, o que foi feito até 1986, quando foram criadas mais seis "linhagens". Desde 1986, são selecionados, anualmente, dez reprodutores (um de cada "linhagem"), com base na primeira lactação de suas mães, desenvolvimento ponderal e tipo. Quanto às fêmeas, todas as novilhas eram incorporadas ao rebanho, desde que não tivessem problemas reprodutivos ou defeitos anatômicos. Ressalta-se que o direcionamento dos acasalamentos sempre foi feito mediante inspeção visual dos livros de escrituração zootécnica.
Os coeficientes de endogamia e valores genéticos de cada indivíduo, para as características produção total de leite (PL), idade ao primeiro parto (IPP), intervalo de partos (IDP) e duração da lactação (DL), foram obtidos por meio do sistema MTDFREML (BOLDMAN et al., 1995). Os registros utilizados, provenientes das cinco primeiras lactações de 1406 vacas do ecótipo Mantiqueira, filhas de 113 reprodutores, foram analisados por meio de modelos (modelo animal) que incluíam, para as características PL, IDP e DL, os efeitos fixos de ano-estação de parto (águas: outubro a março e seca: abril a setembro); como covariável, a idade da vaca ao parto, em meses, com termos linear e quadrático, além dos efeitos aleatórios de animal, de ambiente permanente e erro. Na análise envolvendo a PL, incluiu-se, ainda, o efeito fixo da duração da lactação. No modelo utilizado para o estudo da IPP, considerou-se como efeito fixo o ano-estação de nascimento e como aleatórios, animal e o erro.
Para se verificar o comportamento da endogamia ao longo dos anos, os animais foram agrupados em gerações. A geração de cada animal foi definida de acordo com o coeficiente proposto por BRINKS et al. (1961):
CGI = (CGP + CGM + 2)/2, em que:
CGI = coeficiente de geração do indivíduo;
CGP = coeficiente de geração do pai; e
CGM = coeficiente de geração da mãe.
Foram considerados pertencentes à geração zero (população base) os animais fundadores, os incorporados ao rebanho e os de genealogia desconhecida.
Para se verificar a influência da endogamia sobre os valores genéticos dos animais, para as características reprodutivas e produtivas, foram realizadas análises de regressão. Para isso, os animais foram agrupados em classes de endogamia, conforme descrito na Tabela 1.
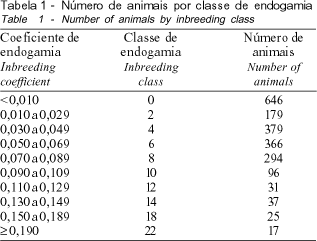
As estatísticas descritivas do coeficiente de endogamia (F) para touros e vacas nascidos entre os anos de 1952 e 1997 encontram-se na Tabela 2. A média de F dos touros foi inferior à das vacas, o que pode ser explicado pela não utilização dos valores genéticos dos machos como critério de seleção. De acordo com MIGLIOR e BURNSIDE (1995), a intensa utilização de touros com valores genéticos mais elevados para produção de leite e de seus componentes tende a aumentar os laços genéticos entre os reprodutores.
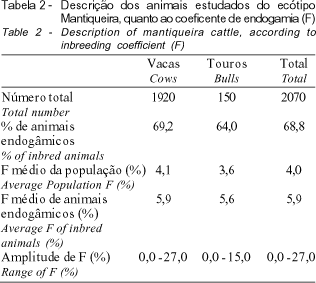
Observa-se ainda, na Tabela 2, que de 1424 animais pertencentes ao rebanho, aproximadamente 69% eram endogâmicos, com coeficiente de endogamia médio de 5,9%. Este valor está próximo a 6,25%, que, segundo FRANCK (1997), é considerado, por muitos autores, como o nível máximo, a partir do qual haveria comprometimento dos desempenhos reprodutivo e produtivo dos animais. De acordo com THOMPSON et al. (2000a), reduções de até 55 kg na produção de leite por lactação estão relacionadas a níveis de endogamia superiores a 7,0%.
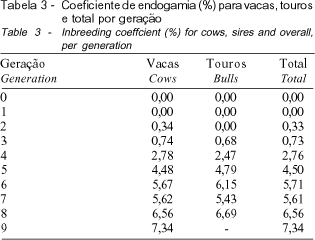
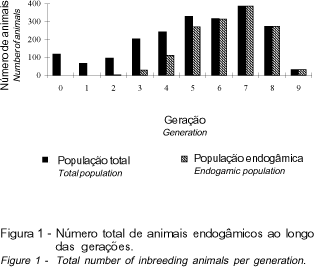
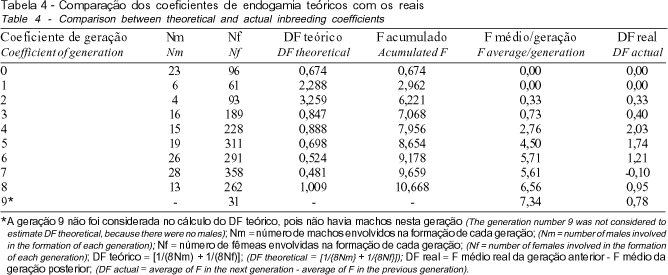
Na Tabela 3, pode ser verificado que houve aumento gradativo da endogamia dos touros e das vacas, ao longo das gerações, fato que, segundo QUEIROZ et al. (2000), pode ser explicado pela própria estrutura dos rebanhos, cujos animais fundadores são pouco aparentados ou de genealogia desconhecida e, com o passar do tempo, mediante a ocorrência de acasalamentos dentro do rebanho, aumenta o grau de parentesco entre os indivíduos, o que leva à endogamia. Na geração 8, o valor médio ultrapassou o limite de 6,25%, chegando a 7,34% na geração 9, indicando que nestas últimas gerações o aumento foi próximo a 1%. A partir da geração 6 (Figura 1), a totalidade dos animais nascidos no rebanho era endogâmica, evidenciando que, a curto prazo, os efeitos da depressão endogâmica podem ser acentuados, ocasionando perda da variabilidade genética, redução da produção de leite bem como da eficiência reprodutiva em geral.
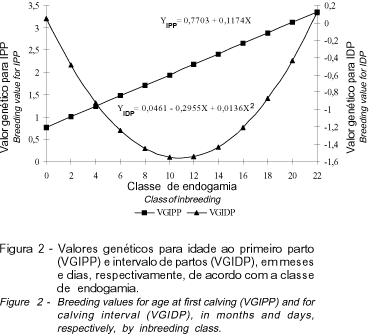
Concordando com os resultados encontrados por PENNA (1990), a partir do nível de 10% de endogamia, houve tendência de aumento nos valores genéticos para IDP (Figura 2), com ponto de mínimo para F=10,86%. Para IPP, observou-se tendência linear de aumento de 0,12 meses nos valores genéticos, para cada acréscimo de 1% no coeficiente de endogamia (Figura 2). Influência significativa da endogamia sobre IPP também foi relatada por THOMPSON et al. (2000a, 2000b), os quais encontraram aumentos de até 17 e 12 dias para IPP, para níveis superiores a 10% de endogamia, nas raças Holandesa e Jersey, respectivamente.
Conforme pode ser verificado na Figura 3, até 10% a endogamia não influenciou nas classificações das vacas quanto ao valor genético para IPP. Todavia, a partir desse nível, os animais com os maiores coeficiente de endogamia eram os que possuíam maiores valores genéticos para IPP, levando-os a ocupar as últimas posições. Tendência semelhante foi observada para IDP.

Na Figura 4 encontram-se as regressões dos valores genéticos para PL e DL, em função das classes de endogamia. Para níveis crescentes de endogamia observa-se tendência linear de diminuição dos valores genéticos para produção de leite. A incrementos de 1% no coeficiente de endogamia correspondem redução de, aproximadamente, 1,15 kg de leite. Efeitos da endogamia sobre a produção de leite também foram constatados por HUDSON e VAN VLECK (1984), SMITH et al. (1998) e THOMPSON et al. (2000a e 2000b). Em relação a DL, tem-se comportamento quadrático dos valores genéticos em relação aos níveis de endogamia, com ponto de máximo para F=11,44%. Até este ponto, houve tendência de aumentos do valores genéticos e, a partir daí, a endogamia influenciou de forma negativa, isto é, reduzindo os valores genéticos dos animais para essa característica. THOMPSON et al. (2000a e 2000b) observaram, para níveis superiores a 5% de endogamia, decréscimo de até 8 e 5 dias, respectivamente, na DL.
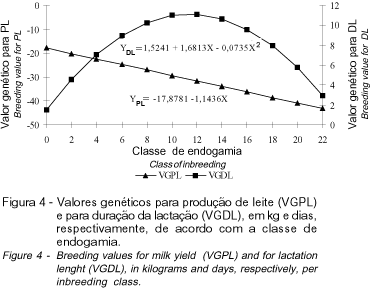
De modo análogo ao observado para IPP, até o nível de 10%, as classificações das vacas quanto ao valor genético para PL não foram influenciadas pela endogamia, entretanto, a partir desse nível, os animais com os maiores coeficiente de endogamia foram aqueles que apresentaram menores valores genéticos para PL (Figura 5). Resultado similar foi encontrado para DL.
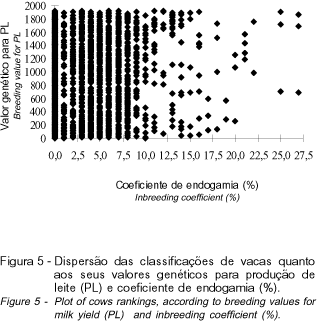
Como o efeito da endogamia sobre os valores genéticos para PL e IPP foi linear, pode-se inferir que os locos que expressam essas características combinam-se aditivamente. Entretanto, para IDP e DL, o comportamento quadrático sugere que a interação epistática entre locos é importante (FALCONER e MACKEY, 1996).
Por meio de um estudo comparativo entre o incremento real (DF real) de endogamia e o incremento esperado, caso os acasalamentos fossem realizados aleatoriamente (DF teórico), concluiu-se que, com exceção das gerações 1 e 2, em que o número de machos é reduzido em relação às demais gerações, o DF teórico se mantém constante, enquanto o DF real aumenta até a quarta geração, diminuindo a partir daí (Tabela 4). Tal comportamento, provavelmente, pode ser explicado pela utilização de um critério de acasalamento de animais não aparentados, iniciado em 1974 e coincidente com o nascimento dos animais pertencentes a geração 5. Destaca-se que, na geração 9, o valor do DF real é, aproximadamente, 1/3 do valor obtido na geração 4, o que demonstra alguma melhoria em relação ao período no qual o parentesco entre os indivíduos, pertencentes à população, não era considerado nos acasalamentos (gerações 0 a 4).
Deve ser considerado que a endogamia influencia as características de crescimento e reprodutivas. Dessa forma, pode-se inferir que a endogamia tenha, possivelmente, maior ação que a verificada neste estudo, uma vez que indivíduos altamente endogâmicos podem ter sido descartados pelo baixo peso ao nascimento, desenvolvimento ponderal inadequado e por problemas reprodutivos e, por isso, não constavam nos registros analisados.
De acordo com MIGLIOR e BURNSIDE (1995), em programas de melhoramento, espera-se o aumento da endogamia em gado de leite, resultante de sua própria estrutura, como por exemplo: alta intensidade de seleção, uso de técnicas reprodutivas, avaliações genéticas mais precisas e uso intenso de alguns reprodutores. Além disso, a seleção com base nos valores genéticos estimados pode aumentar a endogamia, uma vez haverá parentesco entre os animais de maior mérito genético. Especificamente, para o rebanho estudado, tem-se o agravante de não haver introdução de material genético proveniente de outros rebanhos, acelerando o processo de aumento da endogamia.
Ocorreu aumento do coeficiente de endogamia e do número de animais endogâmicos ao longo das gerações, sendo que a partir da sexta geração todos os animais eram endogâmicos.
A endogamia influenciou de maneira negativa os valores genéticos para as características estudadas, promovendo reduções para produção de leite e duração da lactação e aumentos para idade ao primeiro parto e intervalo de partos.
A falta de um programa de acasalamento eficiente e, principalmente, o fato de o rebanho ser fechado têm sido fatores determinantes no crescimento contínuo do nível de endogamia e do número de animais endogâmicos. Assim, a médio prazo, as reduções observadas na fertilidade poderão ser acentuadas.
ALCALÁ, A.M., FRANGANILLO, A.R., CÓRDOBA, M.V. 1995. Analisis genetico de los niveles de consanguinidad en la raza Retinta. Arch. Zootec., 44(166-167):257-265.
BOLDMAN, K.G., KRIESE, L.A., VAN VLECK, L.D. et al. 1995. A manual for use of MTDFREML. A set of programs to obtain estimates of variances and covariances. [DRAFT]. Beltsville: U.S. Department of Agriculture, Agricultural Research Service. 125p.
BRINKS, J.W., CLARK, R.T., RICE, F.J. 1961. Estimation of genetic trends in beef cattle. J. Anim. Sci., 20(4):903.
FALCONER D.S., MACKAY, T.F.C. 1996. Introduction to Quantitative Genetics. 4.ed. New York: Longman Group Limited. 464p.
FRANCK, R. 1997. Inbreeding steals profits. Dairy Herd Management. p.46-50.
GUARAGNA, G.P. Reprodução e endogamia em bovinos Mantiqueira. Ribeirão Preto, SP:FMRP, 1986. 136 p. Tese (Doutorado em Genética) - Faculdade de Medicina de Ribeirão Preto/Universidade de São Paulo,1986.
GUARAGNA, G.P., GAMBINI, L.B., FIGUEIREDO, A.L. 1984. Mantiqueira: 30 anos depois. Nova Odessa: Instituto de Zootecnia, Secretaria de Agricultura e Abastecimento de São Paulo. 30p.
HUDSON, G.F.S., VAN VLECK, L.D. 1984. Effects of inbreeding on milk and fat production, stayability, and calving interval of registered Ayshire catlle in the northeastern United States. J. Dairy Sci., 67(1):171-179.
[ Medline ]
MIGLIOR, F., BURNSIDE, E. B. 1995. Inbreeding of Canadian Holstein cattle. J. Dairy Sci., 78(5):1163-1167.
OLIVEIRA, J.A., BASTOS, J.F.P., TONHATI, H. 1999. Endogamia em um rebanho da raça Guzerá. Rev. bras. zootec., 28(4):721-728.
PENNA, V.M. Efeito da endogamia em características de peso e reprodução da raça Tabapuã. Ribeirão Preto, SP:FMRP, 1990. 102 p. Tese (Doutorado em Genética) - Faculdade de Medicina de Ribeirão Preto/Universidade de São Paulo,1990.
QUEIROZ, S.A., ALBUQUERQUE, L.G., LANZONI, N.A. 2000. Efeito da endogamia sobre características de crescimento de bovinos da raça Gir no Brasil. Rev. bras. zootec., 29(4):1014-1019.
SMITH, L.A., CASSELL, B.G., PEARSON, R.E. 1998. The effects of inbreeding on the lifetime performance of dairy cattle. J. Dairy Sci., 81(10):2729-2737.
[ Medline ]
THOMPSON, J.R., EVERETT, R.W., HAMMERSCHMIDT, N.L. 2000a. Effects of inbreeding on production and survival in Holsteins. J. Dairy Sci., 83(8):1856-1864.
[ Medline ]
THOMPSON, J.R., EVERETT, R.W., WOLFE, C.W. 2000b. Effects of inbreeding on production and survival in Jerseys. J. Dairy Sci., 83(9):2131-2138.
[ Medline ]
WEIGEL, K.A., LIN, W. 2000. Use of computerized mate selection programs to control inbreeding of Holstein and Jersey cattle in the next gneration.
Marcos Vinícius Gualberto Barbosa da Silva /2; 3 William José Ferreira /3, Jaime Araujo Cobuci /3, Guilherme Paes Guaragna /2, Paulo Rogério Palma de Oliveira /2
ferreirawj[arroba]uol.com.br
1. INSTITUTO DE ZOOTECNIA - Parte integrante do projeto IZ - 14-001/52.
2. Pesquisadores Científicos - Núcleo de Pesquisa Zootécnica Geraldo J.R. Alckmin/IZ/APTA/SAA CP 07 - Pindamonhangaba - SP.
3. Alunos de Pós-graduação, DZO/UFV.
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




