 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
Foram conduzidos dois experimentos no conjunto de lisímetros do Laboratório de Biologia Celular e Molecular do CENA-USP em Piracicaba-SP, onde se avaliaram os efeitos dos fatores: cultivar e regime hídrico (irrigação e regime de chuvas), sobre as variáveis: produção de matéria seca (PMS); eficiência do uso da água (EUA); relação haste/folha (H/F); número de hastes (NH); discriminação dos isótopos de carbono (DIC); teor de nitrogênio (N) e nitrogênio total (NT); teor e nitrogênio total derivado da atmosfera (NDA e NTDA). O conjunto de lisímetros foi composto por doze parcelas separadas por paredes de concreto impermeabilizadas de dimensões de 2 m de comprimento, 1,5 m de largura e 2 m de profundidade, sendo 1 m preenchido com brita no1 e 1 m com solo (Latossolo Vermelho-Amarelo distrófico). As sementes de alfafa foram desinfestadas com álcool 700 GL e hipoclorito de sódio (0,5%), para a eliminação de microrganismos contaminantes, conforme mencionado por Araújo & Hungria (1994). Após desinfecção realizou-se a inoculação de Sinorhizobium meliloti SEMIA-116 e peletização com carbonato de cálcio (Somasegaran & Hoben, 1994).
Experimento 1 - A acidez e deficiências nutricionais do solo foram corrigidas com base em análise do solo para fins de fertilidade. A calagem empregada, calculada pelo método da saturação por bases, foi de 1,0 Mg/ha-1 de calcário dolomítico PRNT=85, enquanto a adubação foi de 75 kg.ha-1 de P2O5 na forma de SFS; 30 kg.ha-1 de K2O na forma de KCl, 30 kg.ha-1 de FTE BR-12 e 90 g.ha-1 de molibdênio na forma de molibdato de sódio. Após cada corte foram realizadas adubações de reposição tendo como base o potencial de extração para 2,5 Mg.ha-1 de matéria seca a cada corte. Dessa forma, aplicou-se 60 kg.ha-1 de P2O5 na forma de SFS, 72 kg.ha-1 de K2O na forma de KCl, 30 kg.ha-1 de FTE BR-12 e 50 g.ha-1 de molibdênio na forma de molibdato de sódio.
O experimento foi delineado no esquema inteiramente casualizado em parcelas subdivididas, justificada pela necessidade de se isolar o efeito da irrigação. Foram utilizados oito dos doze lisímetros disponíveis. Dentro de cada lisímetro foram semeadas três diferentes cultivares cada qual em quatro linhas de 1,5 m de comprimento, espaçadas em 0,18 m entre si. Dessa maneira, quatro lisímetros foram mantidos sob irrigação e quatro sob regime de chuva. Foram coletadas duas subamostras em cada subparcela. O experimento teve início no mês 06/98. Foram realizados dois cortes de homogeneização após 60 e 90 dias da germinação, quando então teve início o controle de irrigação. A avaliação ocorreu durante o período de 10/98 a 03/99 por meio de seis cortes realizados sempre que as plantas apresentaram 10% de hastes floridas ou quando os perfilhos basais tivessem de 5 a 8 cm de altura.
Foram utilizados os cultivares Crioula Chilena, Crioula Brasileira, Victoria, UC-Cíbola, Moapa-69, Mecca-I, Mecca-II, WL-516, WL-605, SW-14, Pioneer-5312, Pioneer-XAI32.
Experimento 2 - Dos 12 cultivares iniciais foram selecionados seis em razão do exposto: Crioula Brasileira é o cultivar de maior difusão entre os produtores brasileiros; Crioula Chilena, maior produção de matéria seca em campo; Pioneer XAI32 apresenta boa produção de matéria seca em campo; Mecca tem bom desempenho quanto a FBN; UC-Cíbola tem boa produção de matéria seca em lisímetros. O experimento foi delineado no esquema inteiramente casualizado em parcelas subdivididas. Dentro de cada um dos doze lisímetros foram semeadas duas diferentes cultivares cada qual em seis linhas de 1,5 m de comprimento, espaçadas em 0,18 m entre si. Foram coletadas duas subamostras em cada subparcela. O experimento foi avaliado durante o período de aproximadamente um ano, 07/99 a 04/00, com cortes realizados quando as plantas apresentaram 10% de hastes floridas. Com base na análise química para fins de fertilidade do solo, cada lisímetro recebeu a quantidade de: Lisímetro 5=1,63 Mg.ha-1; L6=1,28 Mg.ha-1; L7= 1,44 Mg.ha-1; L8=1,42 Mg.ha-1; L9=1,32 Mg.ha-1; L10=1,63 Mg.ha-1; L11=1,59 Mg.ha-1 e L12=1,06 Mg.ha-1, de calcário dolomítico PRNT=98, calculada pelo método da saturação por bases. Aplicou-se ainda 78 kg.ha-1de K2O na forma de KCl, 30 kg.ha-1de FTE BR-12 e 90 g.ha-1de molibdênio na forma de molibdato de sódio. Após cada corte foram realizadas adubações repositivas iguais as do Experimento 1.
A capacidade de retenção de água do solo foi previamente determinada para uniformização da umidade, indispensável à avaliação das parcelas mantidas sob condições artificiais. A quantidade de água no solo disponível para a planta (AD), foi determinada com o auxílio da fórmula: AD=[(CC - PMP) . DG] . 300/100, onde: CC=29%; PMP=17,73% e DG=1,22 g.cm-3. Desse modo, a AD calculada foi igual a 41,25 mm. Portanto, toda vez que se observou 0,5 atm nos tensiômetros aplicou-se por asperssão ½ AD ou 20,60 mm.
A altura em cm da coluna de mercúrio correspondente a 0,5 atm indica que cerca da metade da água disponível gasta foi determinada pela equação: T=(12,6hHg hc Hp), em que hHg = altura do mercúrio a partir do nível do reservatório (cm); hc = altura do nível de mercúrio em relação ao solo (cm); hp = profundidade da cápsula no solo (cm). Foram instalados dois tensiômetros com cápsulas a 10 e 20 cm de profundidade, nos quais, a altura média de 43 cm do menísculo em relação à cubeta de mercúrio representou 0,5 atm ou o ponto ideal de início da reposição de água via irrigação.
A cada 28 a 35 dias, o que corresponde a um período necessário para formação, maturação e início do florescimento, as parcelas experimentais foram avaliadas quanto à: produção de matéria seca: realizou-se a colheita de quutro subamostras de 0,2 m2 cortadas de 6 a 8 cm do solo. O material colhido foi seco em estufa com ventilação forçada a 60oC durante 72 horas. Antes, as hastes maduras e juvenis foram contadas fornecendo o número de hastes: relação haste/folha: quatro subamostras de cada cultivar foram coletadas para a estimativa da relação haste/folha, as quais foram secas em estufa com ventilação forçada a 60oC durante 72 horas. Considerou-se haste o caule central e folha, o conjunto de gemas apicais, limbo, pecíolo e flores.
Eficiência do uso da água: obtida pela relação entre a PMS e ETm, que por sua vez, foi obtida pela relação entre o Kc da alfafa e a ETo mensurada em tanque tipo Classe A, instalado no posto agrometereológico da ESALQ/USP em Piracicaba-SP.
Teor de nitrogênio: uma subamostra de cada repetição da matéria seca foi moída e homogeneizada para posterior digestão sulfúrica e análise quanto ao teor de nitrogênio por colorimetria.
O nitrogênio total foi obtido pela relação entre o teor de nitrogênio e produção de matéria seca.
Nitrogênio derivado da atmosfera: a porcentagem de nitrogênio na matéria seca oriunda da fixação biológica de nitrogênio foi obtida com a determinação da composição isotópica por espectrometria de massa.
O nitrogênio total derivado da atmosfera foi obtido pela relação entre o nitrogênio derivado da atmosfera e a produção de matéria seca.
Discriminação dos isótopos de carbono: a composição isotópica foi determinada no Laboratório de Isótopos Estáveis do CENA/USP, com o espectrômetro de massa ANCA-IRMS, conforme Barrie & Prosser (1997). A amostra padrão para controle das análises foi a sacarose de beterraba (d relativo ao PDB = -24.08o/oo). A composição isotópica da atmosfera foi considerada para todos os efeitos como sendo -8o/oo em relação à referência internacional, Pee Dee Belemnite (PDB).
O controle de ervas daninhas foi realizado manualmente, enquanto que o controle de pragas foi realizado em duas ocasiões com o uso do inseticida Deltametrina (0,4 L/ha), para controle de pulgão. Não foi necessário o controle de doenças. A análise estatística dos resultados dos experimentos foi realizada com o software SAS. Avaliou-se a hipótese de igualdade de resposta dos tratamentos pelo teste F de Snedecor e Fisher e, caso essa hipótese fosse rejeitada, os dados foram submetidos ao teste Tukey de comparações múltiplas de médias, para identificação de possíveis diferenças estatísticas entre as médias obtidas nos tratamentos, conforme descrito por Gomes (1987). As análises de correlação, bem como a determinação dos níveis de significância pelo teste T, foram feitas com o programa SANEST.
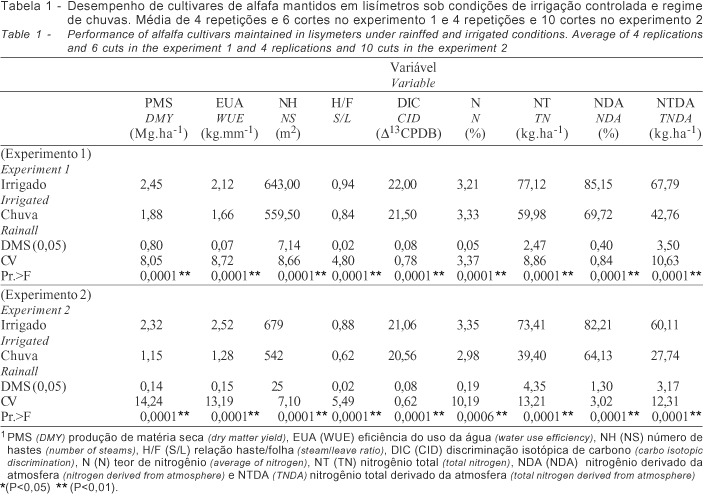
Nos experimentos, a chuva não foi suficiente para garantir níveis satisfatórios de produção e qualidade proporcionando resultados significativamente inferiores (P<0,01) ao tratamento irrigado em todas as avaliações (Tabela 1), o que demonstra a dependência da cultura ao fornecimento constante de água.
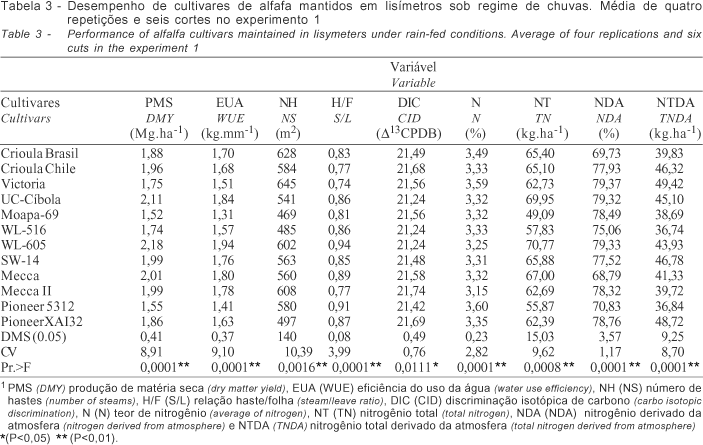
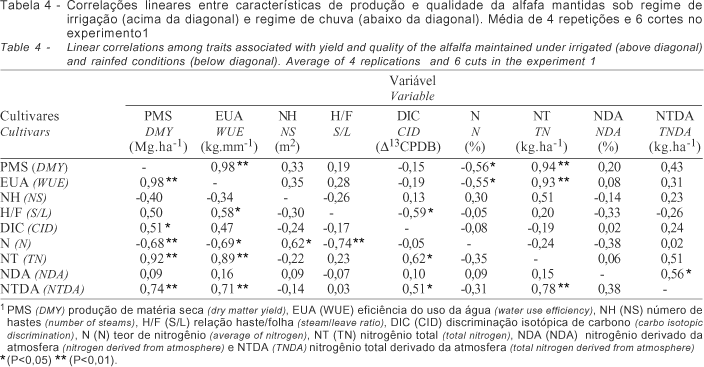
O desempenho dos cultivares dentro de cada regime hídrico no experimento 1 foi diferenciado. Sob regime de chuva (Tabela 1) os cultivares apresentaram significativo aumento de folhas e teor de nitrogênio e redução do nitrogênio total em razão da menor expansão de hastes e produção de matéria seca. No experimento sob condições de irrigação (Tabela 2), alguns cultivares como a Crioula Chilena, UC-Cíbola, WL-605 e Mecca, apresentaram desempenho significativamente superior em relação às características quantitativas avaliadas (P<0,01). O mesmo quadro foi observado também, em condições de chuva (Tabela 3), entretanto em menor escala. A correlação dos dados obtidos (Tabela 4) denota estreita relação entre a produção de matéria seca e a eficiência do uso da água (0,98**) para os dois regimes hídricos demonstrando similaridade no desempenho sob as condições contrastantes. O excesso de chuvas durante o período (10/98 a 03/99) limitou a necessidade de irrigação, influenciando o desempenho dos cultivares. A produção de matéria seca relacionou-se ainda ao N (-0,56*) e NT (0,94**), sob condições de irrigação e a DIC (0,51*), N% (-0,68**), NT (0,92**) e NTDA (0,74**), sob chuva. Não houve correlação entre os dados de eficiência do uso da água e discriminação isotópica de carbono, para os dois regimes hídricos, provavelmente, devido ao curto período de avaliação desse experimento.
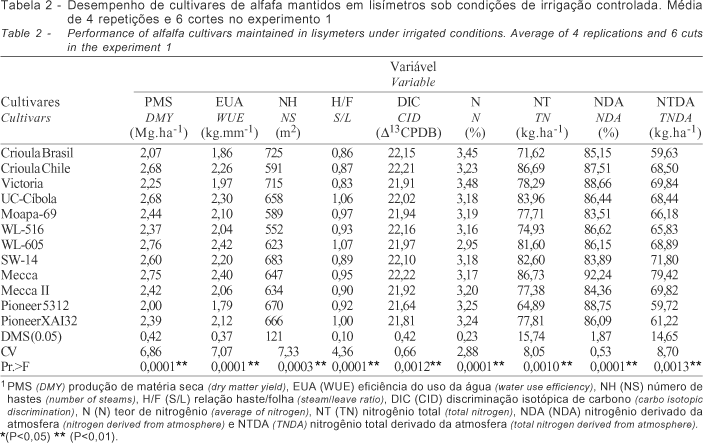
No experimento 2, o desempenho dos cultivares mantidos sob irrigação, quanto à produção de matéria seca (Tabela 5), foi semelhante, sem diferença significativa. Entretanto, sob regime de chuvas os cultivares Crioula Brasileira e Crioula Chilena apresentaram produtividade significativamente superior (P<0,01). Conforme verificado por Oliveira 2001, o menor declínio na produtividade durante os meses de inverno para essas cultivares favoreceu a média anual. O mesmo comportamento é verificado para a EUA. Em relação à DIC o cultivar Mecca apresentou os melhores resultados nos dois regimes hídricos, diferenciando-se dos demais. Em relação ao NTDA é válido apontar o efeito negativo do déficit hídrico sobre a FBN do cultivar Pioneer XAI32. Sob condições de irrigação a PMS correlacionou-se positivamente com a EUA (0,96**), NH (0,80*), NT (0,97**) e NTDA (0,93**) e, negativamente com a DIC (-0,92**). Sob condições de chuva, a PMS correlacionou-se à EUA (0,99**), NT (0,99**) e NTDA (0,81*). Em relação à EUA, no experimento sob irrigação, observa-se correlação negativa com a DIC (-0,94**), conforme descrito na Tabela 6.
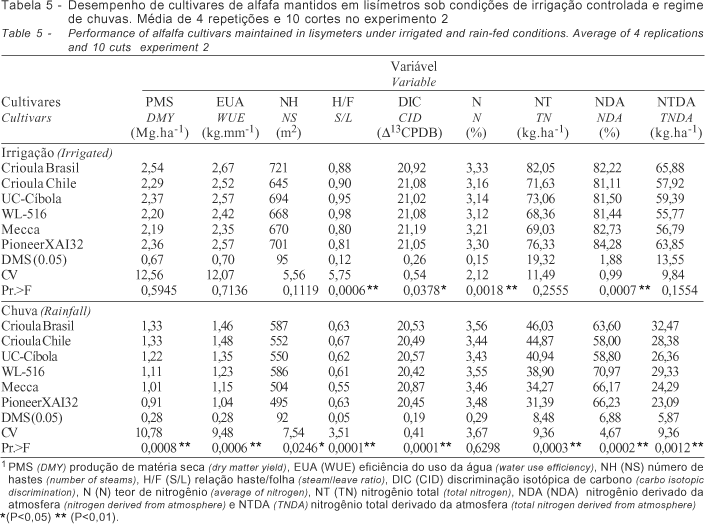
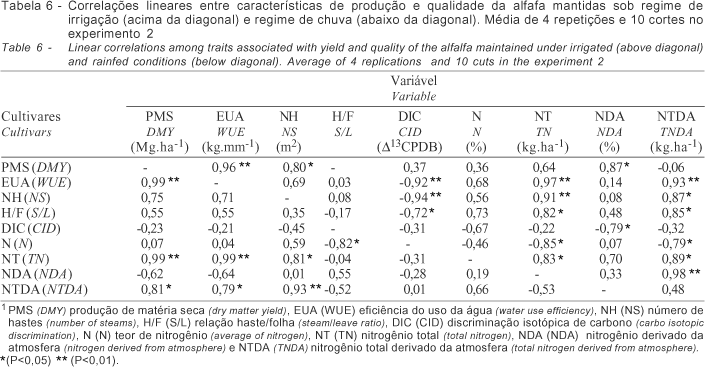
Nesse experimento, a evapotranspiração variou entre 1,7 a 7,1 mm.dia-1 no início e momento do corte, respectivamente. A produção da alfafa relaciona-se diretamente a evapotranspiração da cultura e a água aplicada (Hanson & Orloff, 1998), sendo a eficiência do uso da água a relação entre a água aplicada e a quantidade utilizada ou, ainda, a água necessária aplicada em relação à água utilizada, uma vez que a evapotranspiração da cultura é crescente, conforme descrito por Rassini & Leme (2001) em trabalho com o cultivar Crioula. A determinação da evapotranspiração pelo Kc e Eto estimada em tanque classe A (Doorenbos & Kassan, 1979) tem sido utilizada com sucesso para a cultura da alfafa, conforme descrito por Metochis (1980) e Undersander (1986). Os resultados evidenciaram estreita e positiva relação entre a PMS e EUA independente do ambiente de estudo, resultando em maior produção e eficiência de transpiração. A base fisiológica para variação da DIC em plantas C3 pode ser resumida em termos da razão da concentração do CO2 interno e externo, com tendência de correlação positiva entre a PMS e DIC. Entretanto, essa correlação pode ser negativa em ambientes em que a água seja limitante (Faruquhar & Richards, 1984) ou quando o conjunto de cultivares analisados se mostrem contrastantes (Ehleringer et al., 1991). Segundo Condon et al. (1987) se a correlação positiva entre a DIC e PMS for uma regra, não existe possibilidade de aumento da EUA, uma vez que o aumento da DIC é resultado do aumento da transpiração. Nos experimentos a PMS dos cultivares mantidos sob irrigação correlacionaram-se negativamente com a DIC, resultados semelhantes ao de Johnson & Tieszen (1994) em estudo com 18 cultivares de alfafa. Uma hipótese para justificar os resultados pode estar relacionada aos cultivares estudados que variam quanto à eficiência na utilização da água absorvida no processo de fotossíntese, diminuindo a necessidade de manutenção dos estômatos abertos e resultando em correlação positiva entre a EUA e PMS. Corrobora com essa hipótese a alta concentração de nitrogênio, indispensável à composição das enzimas do sistema fotossintético como a RuBisCO. Outra hipótese seria a análise em separado dos dados obtidos nos experimentos sob irrigação e chuva cuja análise conjunta resultou em correlação positiva invariavelmente (Matus et al., 1995). Nesse experimento, a análise de correlação linear entre PMS e DIC utilizando todo o conjunto de dados resultou em r=0,70, quando avaliado o conjunto de 12 cultivares. Entretanto, quando avaliados os dois conjuntos em separado, observa-se correlação negativa entre a PMS e DIC de r=-0,92**, para plantas mantidas sob irrigação, e r=-0,23 para plantas sob regime de chuvas (Figura 1).
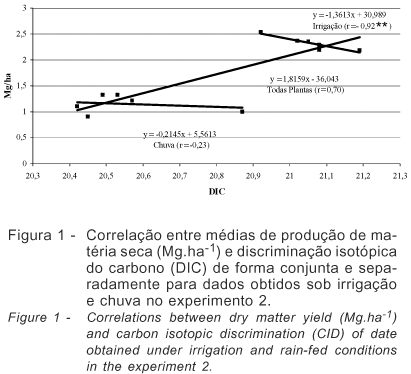
A seleção de plantas para aumento da produtividade com base no aumento da DIC, conforme White et al. (1990), não levou em consideração outras limitações como a FBN e teor de nitrogênio que reflete a concentração de enzimas como a RuBisCO e, segundo o autor essa relação foi inconsistente para plantas de feijoeiro mantidas em diferentes locais.
Para plantas C3, a DIC correlaciona-se negativamente com a EUA (Farquhar et al., 1982) e fatores como sistema radicular e adaptação às condições climáticas são fundamentais (Hubich et al., 1986; White et al., 1990; Johnson & Tieszen, 1994). Nesse estudo, essa correlação foi observada somente quando se restringiu o número de cultivares e estendeu-se o período de avaliação para 12 meses. Para programas de seleção de cultivares Ray et al. (1998) propuseram a identificação de cultivares com maior desenvolvimento radicular, o que facilitaria, sobremaneira, a adaptação ao déficit hídrico, por exemplo. Sob déficit hídrico a relação haste/folha diminui e o teor de proteína bruta aumenta (Oliveira, 1986) aumentando a qualidade e resultando em correlação negativa com a produção (Denment et al., 1986; Robinson et al., 1994). Sob irrigação, o desenvolvimento dos cultivares mais produtivas é otimizado, com menor estacionalidade (Botrel & Alvim, 1997; Rassini & Leme, 2001), devido ao fornecimento de água na época de seca. Essa variação é menor para cultivares que possuem maior capacidade de ajustamento osmótico e manutenção do turgor, mesmo sob baixo potencial hídrico (Wright et al., 1996), como é o caso dos cultivares Crioulas, que possuem ampla adaptação.
O nitrogênio utilizado pela alfafa é uma combinação entre o N absorvido do solo e o NDA obtido via FBN, sendo a alfafa uma planta hábil na utilização tanto da combinação quanto de uma das duas fontes isoladas (Teuber & Phillips, 1986). A diferenciação entre as duas fontes pode ser obtida pela análise do fracionamento isotópico (Shearer & Kohl, 1997). A proporção do N obtido via FBN declina rapidamente quando existe N na solução do solo, mas tende a estabilizar após o segundo corte, devido a alta capacidade de extração de nitrogênio do solo pela alfafa (Hogh-Jensen & Schjoerring, 1997). Durante 3 anos consecutivos de experimentação com alfafa nos pampas da Argentina, Basigalup (2000), encontraram índices de 42% para NDA e 200 kg.ha-1.ano-1 para NTDA. Para a alfafa, 50 a 80% do nitrogênio nas folhas está alocado na forma de proteína fotossintética (Evans, 1989). Parte do nitrogênio (40 a 60%) é transferido para raízes e coroas da alfafa e mobilizado para a emissão e desenvolvimento da parte aérea após o corte. A combinação entre N de reserva e N obtido via fixação tende a variar dentro de um ciclo de produção decrescendo a parcela de reserva para 87, 66 e 52%, após 3, 10 e 30 dias após o corte, respectivamente.
Em linhas gerais, a FBN é positivamente correlacionada a PMS (Heichel et al., 1984) e DIC (Knight et al., 1993), uma vez que existe uma interdependência entre as partes. O metabolismo do N influencia a DIC por estar envolvido na formação da enzima RuBisCO responsável pela assimilação do carbono e principal passo fisiológico na discriminação dos isótopos de carbono em plantas C3. Sob condições de estresse hídrico ou salino o processo pode ser alterado resultando até em relação negativa como obtido por Kumarasinghe et al. (1992) em soja. No experimento 1, os dados são inconsistentes e não descrevem uma tendência, possivelmente por não representarem um ciclo anual. No experimento 2 a condição de estresse hídrico promoveu um desequilíbrio entre as relações, que puderam ser melhor visualizadas na condição de irrigação, em que a correlação positiva entre DIC e FBN foi evidenciada, confirmando a tendência descrita por Knight et al. (1993) para leguminosas eficientes quanto à FBN. Sob condições de irrigação, para uma produção anual de 24,5 Mg.ha-1, a simbiose alfafa/Sinorhizobium meliloti introduziu até 116,69% a mais de nitrogênio a cada corte em relação ao regime de chuvas.
Sob condições controladas a alfafa responde positivamente à irrigação, tanto para a produção de matéria seca quanto para obtenção de nitrogênio e dentre os cultivares estudados, os cultivares Crioula Brasileira e Crioula Chilena apresentam os melhores desempenhos.
A correlação significativa e positiva entre produção de matéria seca e eficiência do uso da água foi observada tanto sob regime de irrigação quanto regime de chuvas, entretanto a correlação entre eficiência do uso da água e discriminação isotópica do carbono foi inconsistente em regime de chuvas e significativamente negativa sob irrigação.
O potencial de fixação biológica do nitrogênio (NDA) correlaciona-se positivamente com a produção de matéria seca para o ciclo de um ano.
À Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP), pelo suporte financeiro.
ARAÚJO, R.S.; HUNGRIA, M. Microrganismos de importância agrícola. Brasília: EMBRAPA, CNPAF, CNPSo, 1994. 236p.
ASSENG, S.; HSIAO, T.C. Canopy CO2 assimilation, energy balance and water use efficiency of an alfalfa crop before and after cutting. Field Crop Research, v.67, p.191-206, 2000.
ATKINS, C.A. Efficiencies and inefficiencies in the legume/Rhizobium symbiosis - a review. Plant and Soil, v.82, p.273-284, 1984.
BASIGALUP, D.H. Alfalfa dinitrogen fixation in Manfredi, Argentina. In: North American Alfalfa Improvement Conference, 37., 2000, madison. Proceedings... Madison: 2000. p.325.
BARRIE, A.; PROSSER, S.J. Automated analysis of light-element stable isotope by ratio mass spectrometry. In: BOUTTON, T.W.; YAMASAKI, S. (Eds.) Mass spectrometry of soils. New York: Marcel Dekker, 1997. p.1-46.
BOTREL, M.A.; ALVIM, M.J. Avaliação de cultivares de alfafa na zona da mata de Minas Gerais. Pesquisa Agropecuária Brasileira, v.32, n.9, p.971-975, 1997.
BOUTTON, T.W. Stable carbon isotope ratios of soil organic matter and their use a indicators of vegetation and climate change. In: BOUTTON, T.W., YAMASAKI, S. (Eds.) Mass spectrometry of soils. New York: Marcel Dekker, 1997. p.47-82.
CONDON, A.G.; RICHARDS, R.A.; FARQUHAR, G.D. Carbon isotope discrimination is positively correlation with grain yield and dry matter production in field-grown wheat. Crop Science, v.27, p.997-1001, 1987.
CUNHA, G.R., PAULA, J.R.F., BERGAMASCHI, H. et al. Evapotranspiração e eficiência no uso da água em alfafa. Revista Brasileira de Agrometereologia, v.2, p.23-27, 1994.
DENMENT, M.W., TEUBER, L.R., BOURQUE, D.P. et al. Changes in forage quality of improved alfalfa populations. Crop Science, v.26, p.1137-1146, 1986.
DOORENBOS, J.; KASSAM, A.M. Efectos del agua sobre el rendimiento de los cultivos. Roma: FAO, 1979. 212p.
EBSON, J.S.; PETROVIC, A.M.; DAWSON, T.E. Relationship between carbon isotope discrimination, water use efficiency and evapotranspiration in Kentucky bluegrass. Crop Science, v.38, p.157-162, 1998.
EHLERINGER, J.; KLASSEN, S.; CLAYTON, C. et al. Carbon isotope discrimination and transpiration efficiency in common bean. Crop Science, v.31, p.1611-1615, 1991.
EVANS, J.R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologie, v.78, p.9-19, 1989.
FARQUHAR, G.D.; O'LEARY, M.H.; BERRY, J.A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Australian Journal of Plant Physiology, v.9, p.121-137, 1982.
FARQUHAR, G.D.; RICHARDS, R.A. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes. Field Crops, v.11, p.539-552, 1984.
FARQUHAR, G.D.; LLOYD, J. Carbon and oxygen isotope effects in the exchange of carbon dioxide between terrestrial plants and atmosphere. In: EHLERINGER, J.R.; HALL, A.E.; FARQUHAR, G.D. (Eds.) Stable isotope and plant carbon-water relations. San Diego: Academic Press, 1993. p.47-70.
FISHBECK, K.A.; HEICHEL, G.H.; VANCE, C.P. Dry matter, nitrogen distribution and nitrogen fixation in contrasting alfalfa symbioses. Crop Science, v.27, p.1205-1209,1987.
GOMES, F.P. A estatística moderna na agropecuária. Piracicaba: Potafós, 1987. 160p.
HALL, A.E.; MUTTER, R.G.; HUBICK, K.T. et al. Genotypic differences in carbon isotope discrimination in cowpea under wet and dry field conditions. Crop Science, v.30, p.300-305, 1990.
HANSON, B.;ORLOFF, S. Double your yield with half the water: Is it possible? In: 1998 CALIFORNIA/NEVADA ALFALFA SYMPOSIUM, 1998, Davis. Proceedings ... Davis: University of California, 1998. p.75-87.
HARDARSON, G.; ZAPATA, F.; DANSO, S.K.A. Field evaluation of symbiotic nitrogen fixation by rhizobial strain using 15N methodology. Plant and Soil, v.82, p.369-375, 1984.
HEICHEL, G.H., HARDARSON, G., BARNES, D.K. et al. Dinitrogen fixation, herbage yield, and rhizobial preference of selected alfalfa clones. Crop Science, v.24, p.1093-1097, 1984.
HOGH-JENSEN, H.; SCHJOERRING, J.K. Effects of drought and inorganic N form on nitrogen fixation and carbon isotope discrimination in Trifolium repens. Plant Physiology Biochemistry, v.35, p.55-62, 1997.
HUBICK, K.T.; FARQUHAR, G.D.; SHORTER, R. Correlation between water-use efficiency and carbon isotope discrimination in diverse peanut (Arachis) germplasm. Australian Journal of Plant Physiology, v.13, p.803-816, 1986.
JASSEN, D.L.; BARNES, D.K.; VANCE, C.P. et al. Variation for activity of nodule nitrogen and carbon assimilating enzymes in alfalfa. Crop Science, v.27, p.627-631, 1994.
JOHNSON, R.C.; TIESZEN, L.L. Variation for water use efficiency in alfalfa germplasm. Crop Science, v.34, p.452-458, 1994.
KNIGHT, J.D.; VERHEES, F.; Van KESSEL, C. et al. Does carbon isotope discrimination correlate with biological nitrogen fixation? Plant Soil, v.153, p.151-153, 1993.
KUMARASINGHE, K.S.; KIRDA, C.; MOHAMED, A. et al. 13C isotope discrimination correlates with biological nitrogen fixation (Glycine max L. merrill). Plant Soil, v.139, p.145-147, 1992.
MARIOTTI, A.; MARIOTTI, F.; AMARGER, N. et al. Fractionnements isotopiques de lázote lors des processus d'absorption des nitrates et de fixation del, azote atmposphérique par les plantes. Physiologie Végetale, v.8, p.163-181, 1980.
MATUS, A.; SLINKARD, A.E.; Van KESSEL, C. Carbon isotope discrimination and indirect selection for seed yield in lentil. Crop Science, v.35, p.679-684, 1995.
METOCHIS, C. Irrigation of lucerne under semi-arid conditions in Cyprus. Irrigation Science, v.1, p.247-252, 1980.
OLIVEIRA, P.R. Avaliação da produção de feno de alfafa (Medicago sativa L.). Piracicaba: Escola Superior de Agricultura "Luiz de Queiroz", 1986. 75p. Dissertação (Mestrado) - Escola Superior de Agricultura "Luiz de Queiroz", 1986.
OLIVEIRA, W.S.; OLIVEIRA, P.P.A.; TSAI, S.M. Associações simbióticas com a microbiota do solo. In: SIMPÓSIO SOBRE MANEJOS DE PASTAGENS, 16., 1999, Piracicaba. Anais ... Piracicaba: Fundação de Estudos Agrários "Luiz de Queiroz", 1999. p.117-132.
OLIVEIRA, W.S. Seleção de cultivares de alfafa (Medicago sativa L.) eficientes em produção e qualidade da biomassa. Piracicaba: Universidade de São Paulo, 2001. 118p. Tese (Doutorado) - Universidade de São Paulo, 2001.
RACCA, R.; BASIGALUP, D.; BRENZONI, E. et al. Alfalfa symbiotic dinitrogen fixation in the Argentine Pampean Region. In: NORTH AMERICAN ALFALFA IMPROVEMENT CONFERENCE, 36., 1998, Bozeman. Proceedings ... Bozeman, Montana, 1998. p.71.
RAY, I.M.; TOWNSEND, M.S.; MUNCY, C.M. Variation for yield, water use efficiency and canopy morphology among nine alfalfa germplasm. Crop Science, v.38, p.1386-1390, 1998.
RASSINI, J.B.; LEME, E.J.A. Water management for establishment of alfalfa (Medicago sativa). In: INTERNATIONAL GRASSLAND CONFERENCE, 19., 2001, Piracicaba. Proceedings ... Piracicaba: Fundação de Estudos Agrários "Luiz de Queiroz", 2001. p.591.
ROBINSON, F.E.; TEUBER, L.R.; GIBBS, L.K. Alfalfa water stress management during summer in Imperial Valley for water conservation. El Centro, CA: Desert Research and Extension Center, 1994. 57p.
SHEARER, G.; KOHL, D.H. Natural abundance of 15N: Fractional contribution of two sources to a common sink and use of isotope discrimination. In: BOUTTON, T.W., YAMASAKI, S. (Eds.) Mass spectrometry of soils. New York: Marcel Dekker, 1997. p.89-125.
SOMASEGARAN, P.; HOBEN, H.J. Handbook for Rhizobia. Methods in legumeRhizobia technology. New York: Springer-Verlag, 1994. 145p.
TEUBER, L.R.; PHILLIPS, D.A. Influences of selection method and nitrogen environment on breeding alfalfa for increased forage yield and quality. Crop Science, v.28, p.599-604, 1986.
UNDERSANDER, D.J. Alfalfa (Medicago sativa L.) growth response to water and temperature. Irrigation Science, v.8, p.23-33, 1986.
WHITE, J.W.; CASTILLO, J.A.; EHLERINGER, J. Associations between productivity, root growth and carbon isotope discrimination in Phaseolus vulgaris under water deficit. Australian Journal of Plant Physiology, v.17, p.189-198, 1990.
WRIGHT, G.C.; WHITFIELD, D.M.; GYLES, O.A. et al. Effects of frequency of irrigation and gypsum treatment on leaf water potential and leaf stomatal conductance of lucerne (Medicago sativa L.) grown on a clay soil. Irrigation Science, v.7, p.73-82, 1996.
ZHU, Y.P.; SHEAFFER, C.C.; RUSSELLE, M.P. et al. Dinitrogen fixation of annual Medicago species. In: NORTH AMERICAN ALFALFA IMPROVEMENT CONFERENCE, 35., 1996, Oklahoma City. Proceedings ... Oklahoma City, 1996. p.75.
Wladecir Salles de OliveiraI; Patrícia Perondi Anchão OliveiraII; Moacyr CorsiIII; Paulo César Ocheuze TrivelinIV; Siu Mui TsaiV
pcotrive[arroba]cena.usp.br
IAluno de Doutorado, CENA/Universidade de São Paulo. E.mail: wsolivei[arroba]carpa.ciagri.usp.br
IIPesquisadora, EMBRAPA/CNPGL - Núcleo Sul. E.mail: ppaolive[arroba]cnpgl.embrapa.br
IIIProfessor Doutor, ESALQ/Universidade de São Paulo. E.mail: moa[arroba]carpa.ciagri.usp.br
IVProfessor Doutor, CENA/Universidade de São Paulo. E.mail: pcotrive[arroba]cena.usp.br
VProfessora Doutora, CENA/Universidade de São Paulo. E.mail: tsai[arroba]cena.usp.br
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




