 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
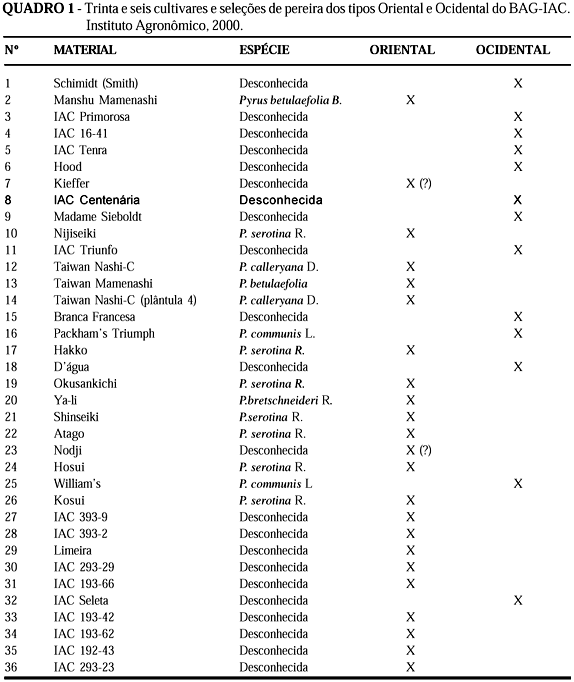
A pêra, pomácea pertencente ao gênero Pyrus, é a terceira fruta mais produzida no mundo depois da uva e maçã. O Brasil produz cerca de 20 mil toneladas anuais de peras das espécies Pyrus communis L. e P. serotina R., porém consome quase dez vezes mais, equivalente a 1,2 kg.pessoa-1. Assim, o mercado brasileiro torna-se bastante dependente de importações, sendo a Argentina, Chile, Estados Unidos, Uruguai e Portugal os principais fornecedores (Oliveira et al., 2000). O gênero Pyrus, que conta com, pelo menos, 22 espécies, é originário de regiões montanhosas da China Ocidental (Vavilov, 1951), sendo que todas as espécies são nativas do continente asiático ou europeu (Rehder, 1967). A domesticação da espécie cultivada, P. communis L., provavelmente, deu-se ao leste da montanha do Cáucaso e Ásia Menor (Vavilov, 1951). Além das pereiras ocidentais (P. communis L.), amplamente cultivadas na Europa e América, também são fartamente encontrados os tipos asiáticos ou orientais (P. serotina R., P. ussuriensis R. e P. bretschneideri R.) na China e Japão (Kikuchi, 1946). Todas as espécies do gênero Pyrus são diplóides (2n = 34, x = 17), preferencialmente alógamas, sendo que existem alguns casos de poliplóides na espécie P. communis L. (Zielinski and Thompson, 1967).
Os programas de melhoramento de peras voltam-se principalmente para a qualidade do fruto, embora os atributos organolépticos possam diferenciar de um país para outro. Esse é o caso das pereiras européias em relação às asiáticas. Os atributos das peras européias, como casca lisa, aromáticas e tipo amanteigado, são menores no oriente, que prefere aquelas mais suculentas, adocicadas e ferruginosas. De modo geral, em ambos os casos, as cultivares com adequada adaptação e resistentes a pragas e doenças somente são aceitáveis se possuírem características desejáveis de fruto. Cruzamentos interespecíficos entre acessos das espécies asiáticas ou orientais, catalogadas no Brasil como Pyrus serotina Rehder, P. betulaefolia Bunge, P. calleryana Decaisne, P. bretschneideri Rehder, podem fornecer características fitotécnicas e organolépticas interessantes. Estas espécies têm, em sua grande maioria, características desejáveis, como fruto doce e crocante, além de rusticidade, ampla adaptação a variados tipos de clima e solo, alta produtividade e importante fonte genética de resistência a várias doenças e pragas que prejudicam as pereiras européias, da espécie P. communis L. No caso especifico do programa de melhoramento genético de pereiras do Instituto Agronômico (IAC), a obtenção de híbridos intra e interespecíficos, representados por pereiras asiáticas e européias, tem sido um dos objetivos, no sentido de se desenvolver cultivares com características agronômicas complementares, ou seja, rusticidade de planta e alta qualidade de frutos.
No entanto, dentro das espécies, os tipos são muito parecidos e, freqüentemente, a classificação é dificultada. Provavelmente, as espécies utilizadas podem ser confundidas com subespécies, variedades botânicas ou híbridos interespecíficos. Desta forma, muitos acessos existentes em bancos de germoplasma de pereira podem apresentar identidade genética desconhecida ou classificação botânica inadequada. Além disso, em muitos casos, as informações morfofenológicas disponíveis são insuficientes para discriminar genótipos aparentados.
Nos últimos anos, com o desenvolvimento da biotecnologia, diversas técnicas de marcadores moleculares têm permitido indicar com precisão as variações genética presentes no DNA de um organismo qualquer. Os marcadores moleculares mais utilizados atualmente, como SSR (single sequence repeat) ou polimorfismo de microssatélite (pequenas seqüências com um a quatro nucleotídeos de comprimento repetidas em tandem) e AFLP (amplified fragment lenght polymophism) ou polimorfismo de comprimento de fragmentos amplificados, requerem mais tecnologia para sua execução. Portanto, apesar da vantagem dos marcadores SSR de possibilitarem maior conteúdo de informação de polimorfismo, devido à expressão co-dominante e o multialelismo, em relação ao RAPD, que tem comportamento dominante, detectando apenas um alelo por loco, sua utilização é limitada pela necessidade prévia do desenvolvimento dos marcadores. Por sua vez, apesar de a análise AFLP possibilitar a obtenção de um grande número de marcadores em uma única análise, oriundos de análise multiplex e combinação do polimorfismo de presença e ausência de sítios de restrição com a ocorrência ou não de amplificação a partir de seqüências arbitrárias, têm comportamento dominante como o RAPD, detectando apenas um alelo por loco. Ainda, por ser uma análise mais sofisticada, demanda mais etapas para sua execução, sendo mais requerida em espécies de plantas cultivadas que apresentam uma baixa taxa de polimorfismo de DNA (Ferreira & Grattapaglia, 1998). Já o RAPD (Random Amplified Polymorphic DNA) tem sido amplamente utilizado em pesquisas de genética de plantas, inclusive de pereiras, devido a sua facilidade de execução. Weeden et al. (1994), utilizando 70 primers de RAPD, encontraram cerca de 220 fragmentos polimórficos para a população segregante de um 'duplo pseudotestcross' em pêra. Oliveira et al. (1999), empregando a técnica RAPD para verificar o relacionamento entre vários cultivares de Pyrus communis, P. pyrifolia e genótipos de P. cordata, P. bourgaeana e P. pyraster, diferenciaram o padrão de pêra ocidental da oriental e encontraram marcadores específicos para alguns genótipos de cultivares. Monte-Corvo et al. (2000), também utilizando marcadores do tipo RAPD, foram capazes de discriminar genótipos de pêra do grupo oriental de ocidental, assim como dos seus híbridos. O uso de marcadores moleculares do tipo RAPD, associado às avaliações fenotípicas do material, pode ser útil em programa de melhoramento genético de pereira, para caracterização de acessos, bem como identificação rápida de seleções interespecíficas oriundas ou não de cruzamentos controlados. Esse é o caso do banco de germoplasma do Instituto Agronômico, que conta com cinco cultivares de pereiras lançadas pela Instituição e outras seleções IAC, oriundas de cruzamentos ou de polinização aberta, cuja origem e identificação genética ainda são incertas (Barbosa et al., 1997).
Neste trabalho, analisaram-se 36 acessos de pereira da coleção de trabalho do IAC, com a técnica RAPD, para se verificar a possibilidade de identificação e caracterização, além do relacionamento genético entre os acessos.
O DNA das pereiras do Quadro 1 foi extraído de folhas jovens, utilizando-se do método com tampão de brometo de hexadeciltrimetilamônio (CTAB). Cerca de 0,5 g de tecido fresco foi triturado com N2 líquido e incubado a 65oC por 20 minutos com 5 ml de tampão de extração (2% CTAB; 1,4M NaCl; 20mM EDTA PH 8,0; 100mM Tris-HCl pH8, 0; 1% PVP 40; 2% mercaptoetanol).
O extrato obtido foi submetido a uma extração com clorofórmio-álcool isoamílico (24:1), e centrifugada. O DNA na fase aquosa foi precipitado com 54% de volume de isopropanol e transferido para tubo eppendorff de 1,5ml para ser redissolvido com 200 µl TE pH 8,0 (10 mM Tris-HCl e 1mM EDTA). A mistura foi, a seguir, digerida com RNase e extraída uma vez com fenol-clorofórmio-álcool isoamílico e uma vez com clorofórmio-álcool isoamílico. O DNA foi precipitado com isopropanol, lavado com etanol 70%, seco e dissolvido em TE pH 8,0 diluído10X. A quantificação e diluição do DNA foram medidas em fluorímetro e as reações de PCR realizadas no termociclador MJ PTC-100 com um volume de 10 µl. Foram utilizados cerca de 25ng de DNA em tampão de reação Tris-HCl 10mM pH 8,3 com 1,5mM MgCl2; 50mM KCl; 80µM de cada deoxinucleotídeo; 0,1% BSA; 0,1% NP-40 e 0,1 a 0,2µl de Taq DNA polimerase ( 5 unidades/µL). A amplificação foi iniciada a 94oC, por 2 minutos, para a desnaturação inicial seguida por 45 ciclos de 15 segundos, a 94oC (desnaturação), 25 segundos, a 34oC (anelamento) e 50 segundos, a 72oC (extensão), mais uma etapa para a completa extensão de todas as cadeias complementares de 5 minutos, a 72oC. Foram utilizados os primers dos kits A, C, D e F da Operon Technologies para as reações de amplificação.
Os dados foram analisados pelo programa NTSYS, versão 1.70 (Rohlf, 1992), para a construção do dendograma e análise de coordenada principal, utilizando-se somente das bandas polimórficas mais consistentes em, pelo menos, duas amplificações para cada primer. O coeficiente de similaridade de Jaccard a/(n-d) foi estimado a partir dos dados de presença de banda (1) ou ausência (0), onde a = número de coincidências positivas (tipo 1-1), n= número total de amostras e d= número de coincidências negativas (tipo 0-0). A partir dos dados de coeficiente de similaridade gerados pelas bandas RAPD, foram feitas as análises dos agrupamentos, gerando o dendograma pelo método UPGMA e a análise de coordenada principal a partir do método de Gower (1966).
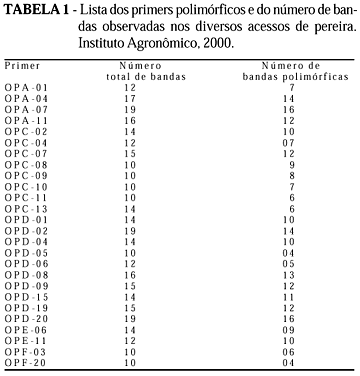
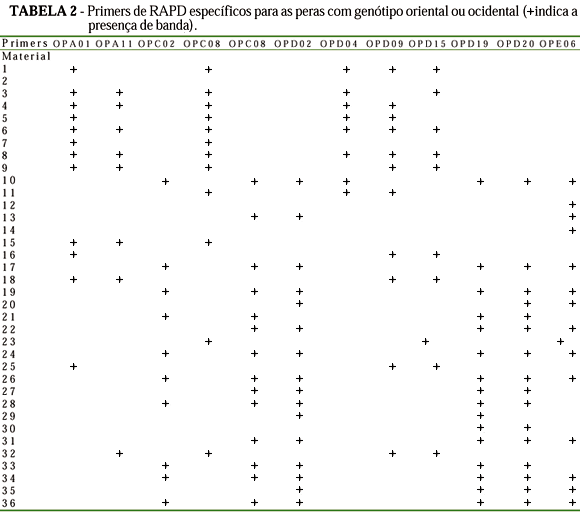
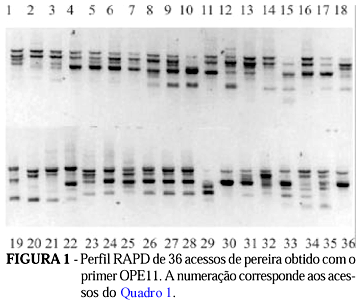
Vinte e seis primers forneceram polimorfismo, com um total de 250 bandas polimórficas, de um total de 353 (Tabela 1). Cada primer originou entre 10 a 19 bandas, sendo evidente o polimorfismo entre os acessos, como mostrado no exemplo de padrão de bandas RAPD obtido com o primer OPE11 (Figura 1). Somente com este primer verifica-se a existência da grande diversidade genética entre os acessos. Pode-se, também, ser vista a semelhança entre os híbridos IAC293-29 (30) e IAC293-23 (36) com a 'Okusankichi' (19), confirmando-a como um dos progenitores, assim como a maior similaridade com as peras orientais, pois o outro progenitor é a pêra 'D'água' (18) que, pelas análises, ficou como sendo do tipo ocidental. Da mesma forma, os híbridos 31 e 33 são mais semelhantes ao progenitor 'Atago' (22), de procedência oriental, do que a 'D'água', o outro progenitor, embora os outros híbridos, 34 e 35, aparentem ser, neste caso, mais próximos de 'D'água' do que 'Atago'. A Tabela 2 apresenta a relação dos primers que diferenciaram as peras com genótipo oriental das ocidentais. Os primers OPC02, OPC08, OPD02, OPD19, OPD20 e OPE06 revelaram bandas específicas para os cultivares orientais números 10; 17; 19; 20; 21; 22; 24; 26; 27; 28; 29; 30; 31; 33; 34; 35 e 36, enquanto os primers OPA01, OPA11, OPC08, OPD04, OPD09 e OPD15 para os ocidentais 1; 3; 4; 5; 6; 8; 9; 15; 16; 18; 25 e 32. A formação dos grupos, utilizando-se do método de agrupamento UPGMA, originou o dendograma da Figura 2. O coeficiente cofonético computado da matrix da árvore mostrou correlação de 0,932 com a matrix de similaridade original, indicando boa correspondência dos agrupamentos aos dados. Verificou-se, pelo dendograma, três principais agrupamentos, ficando distintas as peras ocidentais das orientais e o grupo das Taiwan. No grupo ocidental, reuniram-se, além dos cultivares da espécie P. communis L., 'William's(25) e 'Packham's Triumph' (16), também, as seleções IAC ('P. Triumph' x 'Hood'), como IAC 16-41 (4), 'Triunfo' (11), 'Centenária' (8), 'Primorosa' (3) e 'Seleta' (32), além de 'Tenra' (5) ('P. Triumph' x 'Madame Sieboldt'). Com exceção de Seleta, os demais híbridos IAC formaram um grupo dentro do agrupamento das ocidentais, indicando também grande proximidade genética. 'Hood' e 'Madame Sieboldt' (9), cultivares receptoras e doadoras de pólen, respectivamente, estreitamente próximas, ficaram ligadas com a "Kieffer', dentro do grupo das ocidentais. Ressalte-se que 'Hood' (6) e 'Kieffer' (7) são citados na literatura como supostamente híbridos entre P. communis L. (ocidental) e P. serotina. R. (oriental) (Layne & Quamme, 1975). Ainda dentro do agrupamento das ocidentais, foram relacionadas a D'Água (18), 'Branca Francesa' (15) e, mais distante, a 'Schimidt' (1). Dentro do agrupamento das pereiras asiáticas, ficaram bem definidos dois agrupamentos, sendo um composto pelos acessos da espécie P. serotina R., como 'Nijiseiki' (10), 'Hosui' (24), 'Hakko' (17), 'Okusankichi' (19), Shinseiki (21) e 'Atago' (22). A Shinseiki, suposto híbrido entre 'Nijiseiki' e 'Chojuro', foi confirmada próxima a 'Nijiseiki'. O único acesso da espécie P. serotina R., que não foi relacionado neste agrupamento, foi 'Kosui' (26, oriundo do cruzamento entre 'Kikusui' e 'Wasekojo'), o qual ficou junto às seleções IAC oriundos de polinização livre da D'Água (18): IAC 393-9 (27) e IAC 393-2(28), sugerindo possível ascendência destes híbridos. Ainda dentro do agrupamento dos híbridos entre D'Água e pereiras orientais, o grupo mais próximo, a seguir, foi o formado pelos híbridos IAC 293-29 (30) e IAC 293-23 (36), descendentes de 'Okusankichi' e o IAC 193-66 (31), de 'Atago'. O outro agrupamento próximo foi também formado a partir do cruzamento entre D'Água (18) e 'Atago' (22), constituindo os híbridos IAC 193-42 (33), IAC 192-62 (34) e IAC 192-43 (35). Isso é possível, pois sendo as pereiras 'D'Água' e 'Kosui', vizinhas próximas no lote experimental, podem ter sido interpolinizadas naturalmente. Ainda dentro do grupo das orientais, mais distantes, ficaram a 'Limeira' (29), Nodji' (23) e a 'Ya-li' (20, da espécie P. bretschneideri R.). Finalmente, o terceiro grupo distinto foi formado pelas pereiras porta-enxerto da série Taiwan (12 e 14, da espécie P. calleryana D. e 13, da espécie P. betulaefolia B.) e um pouco mais distante, a 'Manshu Mamenashi' (2, da espécie P. betulaefolia B.). Embora 'Hood', como mencionado anteriormente, seja supostamente híbrido entre P. communis L. e P. serotina R, originou cultivares do cruzamento com a 'P. Triumph' que mostraram maior proximidade genética com as peras ocidentais (Pyrus communis L.), enquanto os híbridos originados entre acessos da P. serotina R. com a 'D'Água' ficaram mais ligadas às orientais, evidenciando maior afinidade da 'Hood' com as pereiras ocidentais. Augustin et al. (1993) encontraram três padrões eletroforéticos da enzima isoperoxidase em acessos da P. calleryana D., indicando que a heterogeneidade constatada nos porta-enxertos é, provavelmente, devido a variabilidade genética. A análise via RAPD confirma a diferença entre duas plantas desta espécie, a Taiwan Nashi-C (12) e Taiwan Nashi-C plântula 4 (14). Apenas com o primer OPE11 já se verificam diferenças entre as plantas, indicando, devido à facilidade da metodologia RAPD, ser este um método prático para se verificar a homogeneidade dos germoplasmas de pereira porta-enxerto. A dispersão gráfica dos dados, obtida pelo método de ordenação de coordenadas principais (Figura 3), originou os mesmos três principais agrupamentos obtidos pelo dendrograma. A primeira (PC1), a segunda (PC2) e a terceira (PC3) coordenadas principais foram responsáveis por 15,15%; 9,64% e 5,04% da variação total, respectivamente.


1. Verificou-se nítida separação entre os genótipos ocidentais e orientais e os das pereiras utilizadas como porta-enxertos.
2. A metodologia RAPD é prática para se identificar genótipo de pereira, podendo ser utilizada para verificar homogeneidade e facilitar a formação de mudas uniformes.
3. A caracterização das cultivares IAC permitiu identificar maior proximidade com a ascendência ocidental que oriental.
4. Para efeito de gestão de recursos genéticos de pereiras, os marcadores RAPD revelam-se importante ferramenta para identificação de duplicatas varietais.
5. Os marcadores RAPD podem auxiliar em muito os melhoristas da pereira, seja para identificação de prováveis parentais de híbridos, cuja origem é desconhecida, seja para estruturar a variabilidade genética no sentido de direcionar cruzamentos entre grupos heteróticos de interesse.
Os autores agradecem aos Técnicos de Apoio Marcilene de Moraes e Antonio Carlos de Carvalho os tratos culturais e a coleta de folhas das pereiras.
Haiko Enok Sawazaki, Wilson Barbosa, Carlos Augusto Colombo
wbarbosa[arroba]iac.sp.gov.br
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




