 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
O fósforo é um mineral de extrema importância para o metabolismo dos animais, pois, além de fazer parte da composição óssea, também participa de muitos processos fisiológicos. Diversas variáveis são utilizadas para determinar a biodisponibilidade desse elemento: ganho de peso, resistência à quebra dos ossos, fósforo no soro, atividade da fosfatase alcalina, fósforo e cinza nos ossos, anormalidades no esqueleto, densidade dos ossos, distribuição do fósforo em tecidos e digestibilidade aparente e verdadeira (Barbosa et al., 1992; Gomes et al., 1992).
Entre as técnicas utilizadas para a determinação da digestibilidade verdadeira, a da diluição isotópica surge como alternativa para a determinação das perdas endógenas fecais e urinárias (Comar et al., 1953; Vitti, 1989). Nos estudos de metabolismo, essa técnica oferece vantagens sobre as demais por apresentar maior rapidez e precisão e ser menos trabalhosa, apesar de não permitir o aproveitamento da carcaça dos animais (Underwood, 1981).
Muitos estudos já foram desenvolvidos com fósforo, e os transtornos observados no desempenho dos suínos, quando se utiliza somente este elemento na forma orgânica como componente da ração, são conhecidos há várias décadas (Chapman et al., 1955). Tal fato ocorre porque a maior parte do fósforo contido nos grãos aparece complexado em uma molécula orgânica chamada de fitato. Como conseqüência, grande parte desse fósforo passa pelo trato digestivo dos animais monogástricos praticamente sem ser digerido, sendo excretado através das fezes, provocando redução no desempenho dos animais e aumentando a poluição ambiental.
Os trabalhos iniciais desenvolvidos com a enzima fitase, apesar de promissores, tiveram como entrave o alto custo no processo de obtenção da enzima. Com o avanço da tecnologia de fermentação a fitase vem sendo comercializada industrialmente, o que tem despertado interesse de nutricionistas preocupados com o alto custo do fósforo inorgânico e com a poluição ambiental (Cromwell, 1991; Lei et al., 1993; Cromwell et al., 1993, 1995a, 1995b).
A enzima fitase atua nas ligações do grupo fosfato, liberando o fósforo e outros minerais que fazem parte dessa molécula (Cromwell, 1991). Além de melhorar a disponibilidade do fósforo, o uso dessa enzima também melhora a disponibilidade de outros minerais, como magnésio, cobre, ferro e zinco.
Cromwell et al. (1993), ao estudarem níveis crescentes de enzima fitase (250, 500 e 1000 UF/g), observaram melhorias lineares no ganho diário de peso e na resistência à quebra dos ossos de suínos. A adição do fósforo inorgânico nas rações de suíno pode ser reduzida ou completamente eliminada pelo uso de 1200 UF/g de fitase na ração, de acordo com os resultados de ganho diário de peso, eficiência alimentar, atividade da fosfatase alcalina no plasma e concentração de cálcio (Lei et al., 1993). Também foi constatada redução de até 50% na excreção do fósforo fecal.
Dentre as diversas fontes de fósforo de origem orgânica, o farelo de arroz desengordurado se destaca pelo elevado percentual em fósforo total, apresentando cerca de 2,13% de fósforo total (Empresa, 1991).
O presente estudo teve por objetivo avaliar rações com milho, farelo de soja e farelo de arroz desengordurado, sem fontes inorgânicas de P, contendo níveis crescentes de fitase, por meio da biodisponibilidade e das perdas endógenas mínimas de P, utilizando a técnica de diluição isotópica e o 32P como traçador metabólico.
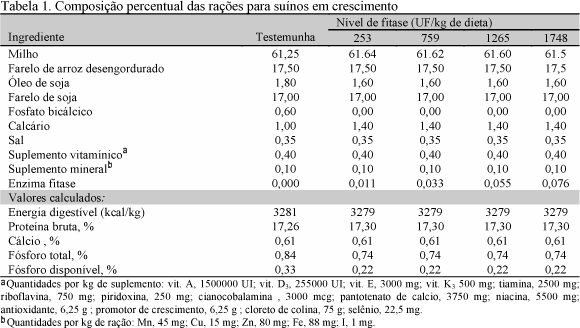
Foram utilizados 20 leitões mestiços, machos castrados, com peso médio de 32,19+2,17kg, alojados em gaiolas metabólicas semelhantes às descritas por Pekas (1968). Os suínos permaneceram por um período de 17 dias nas gaiolas, sendo 10 dias pré-experimentais, para a adaptação às gaiolas e às dietas testes, e sete dias correspondentes à fase de coleta. Os tratamentos consistiram dos níveis de enzima fitase 253, 759, 1265 e 1748 UF/kg de ração, correspondendo a 0,011; 0,033; 0,055 e 0,076 %, respectivamente, como base de cálculo para as rações. A ração testemunha diferiu dos demais tratamentos por ser isenta de enzima fitase e por conter fosfato bicálcico na sua formulação.
O delineamento experimental utilizado foi o de blocos ao acaso, com cinco tratamentos e quatro repetições. Os resultados foram submetidos à análise de regressão polinomial, seguindo os procedimentos do Statistical... (1991). Para comparação das médias dos tratamentos utilizou-se o teste Dunnett.
A enzima fitase (allzyme phytase), produzida a partir de fungos do grupo Aspergillus niger, continha além de atividade de celulase, protease, xylanase e fosfatase ácida, atividade declarada de 2300 unidades de fitase por grama (UF/g). A unidade de fitase, representando a quantidade de enzima que libera 1m mol de fósforo inorgânico do fitato de sódio em um minuto, em condições padrão, foi determinada num substrato de sódio-fitato a 37° C e pH 5,5. A composição percentual das rações de crescimento é apresentada na Tab. 1.
As rações experimentais, fornecidas de acordo com o consumo metabólico (W0,75), foram formuladas à base de milho, farelo de soja, farelo de arroz desengordurado e óleo de soja, enriquecidas com suplemento mineral e vitamínico, de forma a atender às exigências dos suínos na fase de crescimento (National..., 1998; Rostagno et al., 2000), exceto em fósforo disponível. Foram umedecidas na proporção de
1:1 água/ração, e fornecidas em duas refeições diárias, às 7h e 30 min e às 15h e 30 min. Na fase de adaptação, os animais foram estimulados a consumirem o máximo de ração. Na fase de coleta, visando evitar sobras, as quantidades em cada tratamento foram baseadas no menor consumo, dentre os animais de cada bloco, ocorrido no período pré-experimental. A água foi oferecida à vontade.
No primeiro dia da fase experimental cada animal recebeu por via endovenosa 0,5ml de uma solução radioativa com 7,4 MBq de 32P, adquirida no Instituto de Pesquisas Energéticas Nucleares (IPEN). A solução radioativa, preparada de acordo com Vitti (1989), constituiu-se de fosfato de sódio (Na2H32PO4) livre de carregador. Após a aplicação do material radioativo, foram coletados diariamente, durante sete dias, 10ml de sangue em tubos a vácuo heparinizados, em intervalos de 24 horas. As amostras de sangue foram centrifugadas imediatamente após a coleta em 3000rpm durante 10 minutos, separando-se o plasma.
Para a detecção da radioatividade, 0,5ml de plasma foram adicionados em água deionizada em frasco de contagem e a determinação foi obtida por efeito Cerenkov (Nascimento Filho, Lobão, 1977; International..., 1979). Ao volume de 1ml de plasma adicionaram-se 9ml de ácido tricloroacético a 10%, e o teor de fósforo inorgânico determinado por colorimetria (Fiske, Subbarow, 1925).
Amostras de fezes e urina foram coletadas por sete dias. Das fezes 1g foi secado e moído, e após a determinação das cinzas (500 0C), fez-se a digestão com ácido sulfúrico (18N) e determinou-se a radioatividade presente. Para análise do P inorgânico usou-se a digestão das cinzas com ácido clorídrico concentrado, e para sua determinação, o método colorimétrico (Fiske, Subbarow, 1925). A radiotividade da urina foi determinada pelo efeito Cerenkov (Nascimento Filho, Lobão, 1977; International..., 1979) e a do P inorgânico pelo método colorimétrico (Fiske, Subbarow, 1925)
A meia-vida biológica (T1/2) do fósforo foi determinada utilizando-se um sistema cartesiano com atividade específica no plasma, e o tempo, de acordo com a lei do decaimento dos átomos radioativos (International..., 1979). O cálculo foi desenvolvido segundo Lopes (1998), sendo T1/2 = ln2 / K, em que K é a constante de decaimento biológico, calculada pela equação exponencial A = Ao.e-kt, em que A = atividade específica no instante t, Ao = atividade específica inicial (t = o) e t = tempo.
O P endógeno e a biodisponibilidade foram calculados pelas atividades específicas no plasma e nas fezes (Vitti, 1989).
Os resultados relativos ao metabolismo do fósforo em função dos níveis de fitase nas rações são apresentados na Tab. 2.
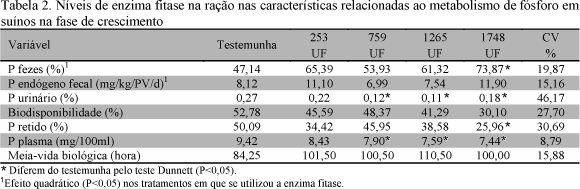
Os níveis crescentes de enzima fitase influenciaram de forma quadrática (P<0,05) a porcentagem de fósforo excretado nas fezes, conforme a equação ![]() = 79,893 0,056X + 301E-7X2, R2 = 0,32, indicando redução da excreção de fósforo até o nível de 931 UF (ponto de mínimo). Este resultado é semelhante aos obtidos por Simons e Versteegh (1990), Cromwell et al. (1995a) e Figueirêdo et al. (2000), que observaram redução da excreção de fósforo com o uso de fitase.
= 79,893 0,056X + 301E-7X2, R2 = 0,32, indicando redução da excreção de fósforo até o nível de 931 UF (ponto de mínimo). Este resultado é semelhante aos obtidos por Simons e Versteegh (1990), Cromwell et al. (1995a) e Figueirêdo et al. (2000), que observaram redução da excreção de fósforo com o uso de fitase.
A enzima fitase atua nos grupos fosfóricos das moléculas de fitato liberando o fósforo e outros minerais para o metabolismo dos animais (Cromwell et al., 1995b). A redução da excreção desse mineral traz benefícios importantes e pode favorecer regiões onde há maior concentração de suínos, ao reduzir a emissão de poluentes para os mananciais de água. Não se observou diferença entre os tratamentos com fitase nos níveis de 253, 759 e 1265 UF e o tratamento testemunha, porém, o nível de 1748 UF apresentou valor superior ao tratamento testemunha (P<0,05), mostrando redução do efeito da fitase.
Os níveis crescentes da enzima fitase influenciaram de forma quadrática (P<0,05) o P endógeno fecal conforme a equação ![]() =1,220,0008X+414E9X2, R2 0,39, indicando redução das perdas endógenas até o nível de 966 UF/kg de ração (ponto de mínimo). Esses valores não foram diferentes dos observados para o tratamento testemunha (P>0,05). A excreção endógena fecal é um mecanismo fisiológico importante no controle da excreção do excesso de fósforo no organismo dos animais (Braithwaite, 1985). Portanto, o efeito sobre essa variável pode indicar que o consumo de P foi suficiente para atender às exigências dos animais, pois em condições de restrição de P, as perdas endógenas fecais são mínimas e iguais à parte obrigatória do metabolismo (Figueirêdo, 1998; Lopes, 1998). Os valores de P endógeno observados neste estudo (11,10, 6,99, 7,54 e 11,90 mgP/kgPV/dia) são superiores aos obtidos por Lopes (1998) (4,75, 4,31, 4,20, 4,09 e 4,0) que considerou essas perdas como obrigatórias. Segundo o autor, as quantidades de fósforo ingeridas não supriram as necessidades dietéticas de P, o que deve ter provocado maior taxa de reabsorção do P endógeno. Não houve diferença entre rações com níveis crescentes de fitase e ração testemunha quanto ao P endógeno (P>0,05), demonstrando que as rações suplementadas com fitase disponibilizaram quantidades de P suficientes para suprirem as exigências dietéticas do mineral. Os valores obtidos para o P endógeno são semelhantes aos da literatura. Jongbloed et al. (1992) observaram que o fósforo endógeno para suínos variou de 5 a 10 mgP/kg PV/dia.
=1,220,0008X+414E9X2, R2 0,39, indicando redução das perdas endógenas até o nível de 966 UF/kg de ração (ponto de mínimo). Esses valores não foram diferentes dos observados para o tratamento testemunha (P>0,05). A excreção endógena fecal é um mecanismo fisiológico importante no controle da excreção do excesso de fósforo no organismo dos animais (Braithwaite, 1985). Portanto, o efeito sobre essa variável pode indicar que o consumo de P foi suficiente para atender às exigências dos animais, pois em condições de restrição de P, as perdas endógenas fecais são mínimas e iguais à parte obrigatória do metabolismo (Figueirêdo, 1998; Lopes, 1998). Os valores de P endógeno observados neste estudo (11,10, 6,99, 7,54 e 11,90 mgP/kgPV/dia) são superiores aos obtidos por Lopes (1998) (4,75, 4,31, 4,20, 4,09 e 4,0) que considerou essas perdas como obrigatórias. Segundo o autor, as quantidades de fósforo ingeridas não supriram as necessidades dietéticas de P, o que deve ter provocado maior taxa de reabsorção do P endógeno. Não houve diferença entre rações com níveis crescentes de fitase e ração testemunha quanto ao P endógeno (P>0,05), demonstrando que as rações suplementadas com fitase disponibilizaram quantidades de P suficientes para suprirem as exigências dietéticas do mineral. Os valores obtidos para o P endógeno são semelhantes aos da literatura. Jongbloed et al. (1992) observaram que o fósforo endógeno para suínos variou de 5 a 10 mgP/kg PV/dia.
O P excretado na urina não foi influenciado pelos níveis de fitase (P>0,05). Pelo teste Dunnett verificaram-se valores menores para os níveis 759, 1265 e 1748 UF em relação ao da ração testemunha, comportamento semelhante ao observado para o P no plasma. O excesso de P no plasma pode ser detectado pelas excreções endógena e urinária, dessa maneira, o nível 253 UF apresentou maior excreção do que os outros níveis. Os resultados deste estudo diferem dos obtidos por Figueirêdo et al. (2000), que não observaram efeito da enzima fitase sobre essa variável, ao trabalharem com rações com farelo de arroz integral e enzima fitase para suínos na fase de crescimento. Lopes (1998) pesquisou os efeitos de níveis crescentes de fósforo no metabolismo de suínos e também não observou efeito sobre essa variável. Ao contrário deste experimento, nos trabalhos de Lopes (1998) e Figueirêdo et al. (2000) os animais consumiram quantidades de fósforo total insuficientes para suprirem as suas exigências segundo o National... (1998). Os resultados obtidos nesta pesquisa indicam que as excreções endógenas são mais sensíveis do que a por via urinária para se detectar o excesso de fósforo no metabolismo, mas ainda assim o seu uso é importante.
Para a variável biodisponibilidade não se observou efeito dos tratamentos. O nível 759 UF apresentou valor mais próximo ao da ração testemunha. Segundo Breves e Schroder (1991), os animais monogástricos absorvem o fósforo no intestino delgado de duas maneiras: por um mecanismo ativo saturável e um passivo não saturável. Em níveis altos de consumo de fósforo também ocorre absorção passiva, provocada pela pressão do P exercida nos sítios de absorção (Lopes, 1998). Dessa maneira, para se detectar o efeito é necessário que as diferenças sejam grandes, o que não ocorreu neste estudo.
Os níveis crescentes de fitase não influenciaram a variável P retida. Pelo teste Dunnett, o nível 1748 UF apresentou valor inferior (P<0,05) ao da ração testemunha.
A concentração de P no plasma não foi afetada pelos níveis de fitase na ração (P>0,05). Os valores do tratamento testemunha e do nível 253 UF foram semelhantes, indicando que a fitase foi mais eficaz em disponibilizar o P orgânico nesse nível. O valor médio de P no plasma deste experimento, 8 mg/dl, é considerado normal segundo Gurtler et al. (1984) (variação de 8 a 10 mg/dl).
Os níveis plasmáticos de fósforo e cálcio são regulados pelo hormônio da paratireóide (PTH) e pela calcitonina. O PTH além de aumentar o cálcio e reduzir o fósforo plasmático, aumenta a excreção do fosfato urinário, enquanto que a calcitonina exerce efeito antagônico, inibindo a reabsorção óssea do cálcio e fósforo (Ganong, 1977). A vitamina D também tem participação importante no metabolismo do cálcio e do fósforo, aumentando a absorção intestinal. Assim, em função desses mecanismos fisiológicos, a variável fósforo no plasma torna-se um indicador pobre (Vipperman et al., 1974) para se analisar o teor de fósforo das dietas, principalmente para ensaios de metabolismo, devido à sua curta duração.
Os valores da meia-vida biológica (T½) do 32P no plasma: 101,50; 100,50; 110,50 e 100,00 horas, não foram afetados pelos tratamentos (P>0,05). Os resultados obtidos nesta pesquisa são semelhantes aos obtidos por Lopes (1998) que, ao estudarem os efeitos de níveis crescentes de P consumido no metabolismo de P em suínos, observou valores de 89,03; 100,26; 90,00; 115,98 e 111,94 horas. A meia-vida biológica funciona como indicador do tempo de reciclagem desse elemento no organismo animal. Dessa forma, valor menor de T1/2 significa melhor disponibilidade do mineral na dieta, caracterizado por maior absorção e incorporação nos tecidos e menor perda endógena e fecal.
A variável perda endógena de fósforo foi mais precisa do que a variável biodisponibilidade para avaliar os efeitos da enzima fitase. O melhor aproveitamento do fósforo fítico ocorre no nível 759 UF.
J. A. MoreiraI; D. M. S. S. VittiI; J. B. LopesII; M. A. Trindade NetoIII
messiasn[arroba]usp.br
IUniversidade de São Paulo Centro de Energia Nuclear na Agricultura (CENA) Av. Centenário, 303 - Caixa Postal 96 13400-000 - Piracicaba, SP
IIUniversidade Federal do PiauíCampus Universitário de SocopoTeresina, PI
III Faculdade de Medicina Veterinária e Zootecnia USP-Pirassununga
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




