 Página anterior Página anterior | Voltar ao início do trabalho | Página seguinte  |
Foram utilizados cinco acessos de feijão-de-vagem (UENF 1429, UENF 1432, UENF 1442, UENF 1445 e UENF 1448), identificados com base na sua divergência genética, quantificada por Abreu (2001), e seus 10 híbridos F1 sem inclusão de recíprocos.
Para a realização dos cruzamentos, foram efetuadas quatro semeaduras, sendo que as duas primeiras ocorreram em maio de 2001, com o período de cruzamentos tendo ocorrido entre junho e julho de 2001, totalizando 581 cruzamentos. As outras duas semeaduras foram realizadas nos meses de julho e agosto de 2001, sendo os cruzamentos efetuados entre setembro e outubro de 2001, perfazendo um total de 528 cruzamentos. Na realização dos cruzamentos, os acessos UENF 1429, UENF 1432, UENF 1442 e UENF 1445 participaram como doadores de pólen, e os acessos UENF 1432, UENF 1442, UENF 1445 e UENF 1448, como receptores de pólen. Em todos os cultivos, o acesso UENF 1448, por ser precoce, foi semeado uma semana após os demais genitores. A semeadura foi realizada em julho de 2002, avaliando-se 15 plantas de cada genitor e F1, em casa de vegetação localizada na Universidade Estadual do Norte Fluminense Darcy Ribeiro (UENF), em vasos plásticos com volume de 5,0 dm3 de substrato, contendo 40% de solo, 40% de esterco bovino e 20% de areia; cada vaso contendo uma planta.
Na análise estatística, identificados os blocos em que os pretensos híbridos eram de fato autofecundações dos genitores, com base em marcadores RAPD, procedeu-se à análise de variância para blocos incompletos, utilizando os recursos computacionais do programa SAS (1985).
Na análise dialélica empregou-se o Modelo de Griffing (1956) para dados desbalanceados, utilizando o procedimento proposto por Martins Filho et al. (1992), por meio dos recursos computacionais do programa GENES (Cruz, 2001). Para tanto foram utilizadas as médias reais de cada parental e das combinações dialélicas
Foram avaliados: a) número médio de vagens por planta (NMV), obtido pela razão entre o total de vagens, por tratamento, de todos os blocos em relação ao número de repetições; b) peso médio de vagens por planta (PMV), expresso em g, obtido pela relação entre o peso das vagens de todas as plantas de cada tratamento, em todos os blocos, e o número total de repetições; c) número médio de sementes por vagem (NS), obtido pela contagem do número de sementes em uma amostra de dez vagens por planta, para cada tratamento, em relação ao número de repetições do experimento; d) altura média da inserção da primeira vagem (APV), expressa em cm, obtida pela medição da distância do colo até a inserção da primeira vagem, de todas as plantas de cada tratamento, relativo ao número total de repetições do experimento; e e) comprimento médio da vagem (CV), expresso em cm, obtido pela quantificação do comprimento longitudinal de uma amostra de dez vagens por planta, de todas as plantas de cada tratamento, em relação ao número de repetições.
Para a totalidade das características avaliadas foram registradas diferenças significativas em 1% de probabilidade, o que pressupõe a existência de diferenças genéticas entre os acessos utilizados nos cruzamentos dialélicos, o que é alento quanto à possibilidade de ganhos genéticos no desenvolvimento de futuros trabalhos de melhoramento genético com as populações.
O coeficiente de variação experimental variou de 6,08% para CV a 51,79%, para APV. Embora apenas as características NMV e APV tenham revelado magnitudes percentuais de CV elevadas (Pimentel-Gomes e Garcia, 2002), provavelmente, isso decorreu da unidade experimental ter sido a 'planta', a despeito do experimento ser realizado em blocos em casa de vegetação.
Os valores dos quadrados médios para capacidade geral de combinação foram altamente significativos, em nível de 1% de probabilidade, pelo teste F, para todas as características em questão. No entanto, para capacidade específica de combinação, as características NS e APV não exibiram significância. As significâncias registradas na grande maioria das características indicam a existência de variabilidade, resultante da ação de efeitos gênicos aditivos e não-aditivos, denotando a possibilidade de obtenção de novas cultivares fixadas ou híbridos.
É importante destacar que trabalhos com análise dialélica em feijão-de-vagem são raros, havendo, dentre os poucos estudos, divergência quanto aos resultados obtidos. Rodrigues et al. (1998), por exemplo, na análise dos efeitos gênicos atuantes em dialelo envolvendo cinco genitores de hábito determinado, detectaram que os efeitos de dominância foram preponderantes para altura de planta. Por sua vez, Leal et al. (1979), utilizaram a potencialidade do dialelo parcial em feijão-de-vagem, para investigar os efeitos gênicos sobre características de produção, pela comparação entre a performance de gerações F5 e F6, em relação a F1 e F2, tendo concluído que a aditividade, e também a epistasia, foram predominantes, apesar de haver sido detectadas diferenças entre os genitores dependendo da característica e do ano avaliado. Não obstante, Carvalho et al. (1999), no intuito de avaliar os efeitos gênicos atuantes em oito caracteres agronômicos, utilizaram as cultivares Alessa, Andra, Cota e Cascade, como progenitores em esquema de cruzamento dialélico sem a inclusão de recíprocos. Constatou-se a ocorrência de significância em todas as características para capacidade específica de combinação, indicando o envolvimento de efeito epistático e, ou de dominância no controle gênico.
Na Tabela 1 encontram-se as estimativas dos efeitos da capacidade geral de combinação (gi) de cada acesso para todas características avaliadas. Verifica-se que para a característica NMV, somente os acessos UENF 1429 e UENF 1448 acusaram valores positivos de gi, o que permite antever que, nos cruzamentos em que estes dois genitores participarem, haverá contribuição para o aumento do número médio de vagens. Por outro lado, o genitor UENF 1445, por apresentar alto valor negativo de gi, tende a contribuir indesejavelmente para aumentos em NMV, quando em combinações híbridas.
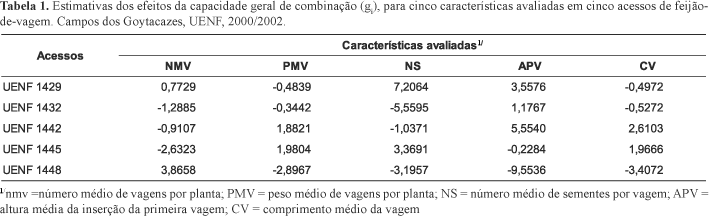
Para a característica PMV, obtiveram-se valores positivos de gi entre os acessos UENF 1445 e UENF 1442, respectivamente, 1,9804 e 1,8821, o que permite a indicação destes em cruzamentos com o objetivo de obter segregantes e linhas puras que promovam o aumento do peso médio de vagens. Porém, os acessos UENF 1448, UENF 1429 e UENF 1432, por revelarem valores negativos de gi, são indesejáveis para proporcionar aumentos no peso médio de vagens.
No que se refere à característica NS, detecta-se que os acessos UENF 1429 e UENF 1445, apresentaram valores positivos de gi, sendo, respectivamente, 7,2064 e 3,3691; portanto, estes genitores tendem a proporcionar híbridos e, conseqüentemente, linhagens com maior número de sementes por vagem, contudo, os genitores UENF 1432, UENF 1448 e UENF 1442, contribuiriam para a redução na magnitude da característica, devido aos valores de gi serem negativos.
Considerando a característica APV, os acessos UENF 1429, UENF 1432 e UENF 1442 expressaram valores de gi positivos, por conseguinte, tendem a contribuir para aumentar a altura de inserção da primeira vagem em indivíduos provenientes de hibridações. De outro modo, apresentaram-se os genitores UENF 1448 e UENF 1445, que tendem a contribuir para a redução na altura de inserção da primeira vagem em programas envolvendo hibridações.
Os acessos UENF 1448, UENF 1432 e UENF 1429, revelaram valores negativos de gi, contribuindo para redução do comprimento de vagens nos híbridos provenientes de seus cruzamentos. Ao contrário, os genitores UENF 1442 e UENF 1445 revelaram valores positivos de gi para a característica CV, portanto, nos programas que têm por objetivo aumentar o comprimento de vagens, tais acessos contribuirão favoravelmente.
Em decorrência dos resultados, pode-se concluir que os acessos UENF 1429, UENF 1445 e UENF 1442 representam os genitores indicados aos programas de melhoramento que visam à obtenção de híbridos para a seleção de linhagens superiores em gerações avançadas, destacando-se dos demais acessos quanto à característica peso médio de vagens, sendo que o primeiro ainda destacou-se para número médio de vagens, número médio de sementes por vagem e altura média de inserção da primeira vagem; os dois últimos revelaram importância para peso e comprimento médios de vagens.
Com relação à discriminação híbrida, Cruz e Regazzi (2001) preconizam que as melhores combinações devem ser aquelas com maior sij, cujos genitores apresentem alta CGC. Para NMV, as combinações que se destacaram foram, respectivamente, 3x5 (5,5617), 1x4 (4,4794) e 4X5 (4,0977) conforme Tabela 2, o que demonstra que os acessos UENF 1429 e UENF1448, destacados pelos valores positivos de gi (Tabela 1), realmente podem gerar indivíduos superiores para número médio de vagens, vez que, pelo menos um dos acessos participa das combinações identificadas como superiores. Entretanto, a combinação entre ambos os acessos revelou reduzido valor de sij (Tabela 2). Cruz e Vencovsky (1989), comparando alguns métodos de análise dialélica, revelaram o fato de que nem sempre dois genitores de alta capacidade geral, quando cruzados, originam o melhor híbrido do dialelo.

Com relação ao peso médio de vagens, as melhores combinações, de acordo com a Tabela 2, com valores positivos de sij foram 1x5 (0,5794), 2x4 (0,6248), 2x5 (0,6952) e 1x4 (0,3956). Somente os acessos UENF 1442 e UENF 1445 apresentaram valores positivos de gi (Tabela 1), demonstrando que o desempenho dos híbridos foi melhor em relação ao esperado com base na CGC dos genitores.
As combinações 1x2 (4,8124), 1x4 (6,0958) e 4x5 (4,4980) apresentaram-se superiores em relação às demais, para a característica NS (Tabela 2). Os acessos que apresentaram valores positivos de gi (Tabela 1) foram UENF 1429 e UENF 1445, indicando que os híbridos F1 contiveram valores esperados com base na CGC dos genitores.
As combinações 1x2 (6,0530), 1X4 (2,6401), 2x3 (2,5451) e 2x4 (3,5010) destacaram-se para a característica APV (Tabela 2). Os acessos UENF 1429, UENF 1432 e UENF 1442 revelaram valores positivos da CGC para a característica APV (Tabela 1). Por outro lado, foi observado que para a combinação 1x3 (-2,9492), Tabela 2, os genitores não demonstraram satisfatório efeito de complementação gênica.
Ainda pela Tabela 2, para a característica CV, as combinações que expressaram as maiores estimativas positivas de CEC foram: 1x4 (1,4567), 1x5 (0,8184), 2x4 (0,6372) e 2x5 (0,6111), sendo, pois, de interesse para incrementos no comprimento da vagem.
Analisando-se em conjunto as características avaliadas, conclui-se que as combinações 1x2 (UENF 1429 x UENF 1432), 1x4 (UENF 1429 x UENF 1445) 2x4 (UENF 1432 x UENF 1445) e 4x5 (UENF 1445 x UENF 1448) têm alto potencial para geração de materiais superiores.
Agradecimentos
Os autores agradecem a UENF/FAPERJ pela bolsa de mestrado concedida a Marlon Peres da Silva; à Técnica Agrícola do Setor de Melhoramento de Plantas do Laboratório de Melhoramento Genético Vegetal da UENF, Elizabeth Frota Morenz, pelo auxílio na realização dos cruzamentos; e ao Prof. Derly José Henriques da Silva, da UFV, pela cessão dos acessos utilizados nesse trabalho.
ABREU, F.B. Aplicação de técnicas de análise multivariada em acessos de feijão-de-vagem (Phaseolus vulgaris L.) de crescimento indeterminado do banco de germoplasma da UENF. 2001. 69 p. (Tese mestrado), UENF, Campos dos Goytacazes.
CARVALHO, A.C.P.P.; LEAL, N.R.; RODRIGUES, R.; COSTA, F.A. Capacidade de combinação para oito caracteres agronômicos em cultivares rasteiras de feijão-de-vagem. Horticultura Brasileira, Brasília, v.17, n.2, p.102-105, 1999.
COSTA, J.G.; ZIMMERMANN, M.J.O. Melhoramento genético do Feijoeiro. In: ZIMMERMANN, M.J.O.; ROCHA, M.; YAMADA, T. (Ed.) Cultura do feijoeiro: fatores que afetam a produtividade. Piracicaba: POTAFOS, 1988. p.229-248.
CRUZ, C.D. Programa genes: aplicativo computacional em genética e estatística. Viçosa: Editora UFV, 2001. 394 p.
CRUZ, C.D.; REGAZZI, A.J. Modelos biométricos aplicados ao melhoramento genético. Viçosa: Editora UFV, 2001. 390 p.
CRUZ, C.D.; VENCOVSKY, R. (1989) Comparação de alguns métodos de análise dialélica. Revista Brasileira de Genética, v.12, n.2, p.425438.
GRIFFING, B. Concept of general and specific combining ability in relation to diallel crossing systems. Australian Journal Biological Science. v.9, p.463-493. 1956.
LEAL, N.R.; HAMAD, I.; BLISS, F. Combining ability estimates for snap bean traits using early and advanced generations. HortScience. v.14, n.3, p.405-405, 1979.
MALUF, W.R.; BARBOSA, M.L.; RESENDE, M.R.R.; COSTA, H.S.C. A Cultura do feijão de vagem. Disponível em: <http://www2.ufla.br/~wrmaluf/bth065/bth065.html>. Acesso em: 08 nov. 2002.
MARTINS FILHO, S.; CRUZ, C.D.; SEDIYAMA, C.S. (1992) Analysis of Unbalanced Diallel Crosses. Revista Brasileira de Genética, Ribeirão Preto, v.15, n.4, p.853869.
PIMENTEL-GOMES, F.; GARCIA, C.H. Estatística aplicada a experimentos agronômicos e florestais Exposição com exemplos e orientações para uso de aplicativos. Biblioteca de ciências agrárias Luiz de Queiroz. Piracicaba: FEALQ, 2002. v.1, 309 p.
RODRIGUES, R.; LEAL, N.R.; PEREIRA, M.G. Análise dialélica de seis características agronômicas em Phaseolus vulgaris L. Bragantia, Campinas, v.57, n.2, p.241-250. 1998. www.scielo.br
SANTOS, M.L.; BRAGA, M.J. Feijão: aspectos econômicos. In: VIEIRA, C.; PAULA JÚNIOR, T.J.; BORÉM, A. (Ed.) Feijão: aspectos gerais e cultura no Estado de Minas Gerais. Viçosa: Editora UFV, 1998. p.19-53.
SANTOS, F.F.; MATOS, M.J.L.F.; MELO, M.F.; LANA, M.M.; LUENGO, R.F.A.; TAVARES, S.A. Feijão-de-vagem. Disponível em: <http://www.emater.df.gov.br/ecndicasind.html#>. Acesso em: 06 ago. 2002.
SAS Institute Inc. 1985. SAS user's guide: statistics. 5. ed. NC Cary. 958 p.
VIEIRA, C. Perspectiva da cultura do feijão e de outras leguminosas de grãos no país e no mundo. In: ZIMMERMANN, M.J.O.; ROCHA, M.; YAMADA, T. (Ed.) Cultura do feijoeiro: fatores que afetam a produtividade. Piracicaba: POTAFOS, 1988. p.02-19.
Marlon P. da SilvaI; Antônio T. Amaral JúniorI; Rosana RodriguesI; Rogério F. DaherI; Nilton R. LealI; Adilson Ricken SchuelterII - rogdaher[arroba]uenf.br
IUENF, CCTA, Av. Alberto Lamego 2000, Parque Califórnia, 28013-600 Campos dos GoytacazesRJIIUniversidade Paranaense, Campus Toledo, Toledo-PR.
| Página anterior | Voltar ao início do trabalho | Página seguinte |
|
|
|




